No β-N-Methylamino-L-alanine (BMAA) Was Detected in Stranded Cetaceans from Galicia (North-West Spain)

 , , and
, , and 
Abstract
:1. Introduction
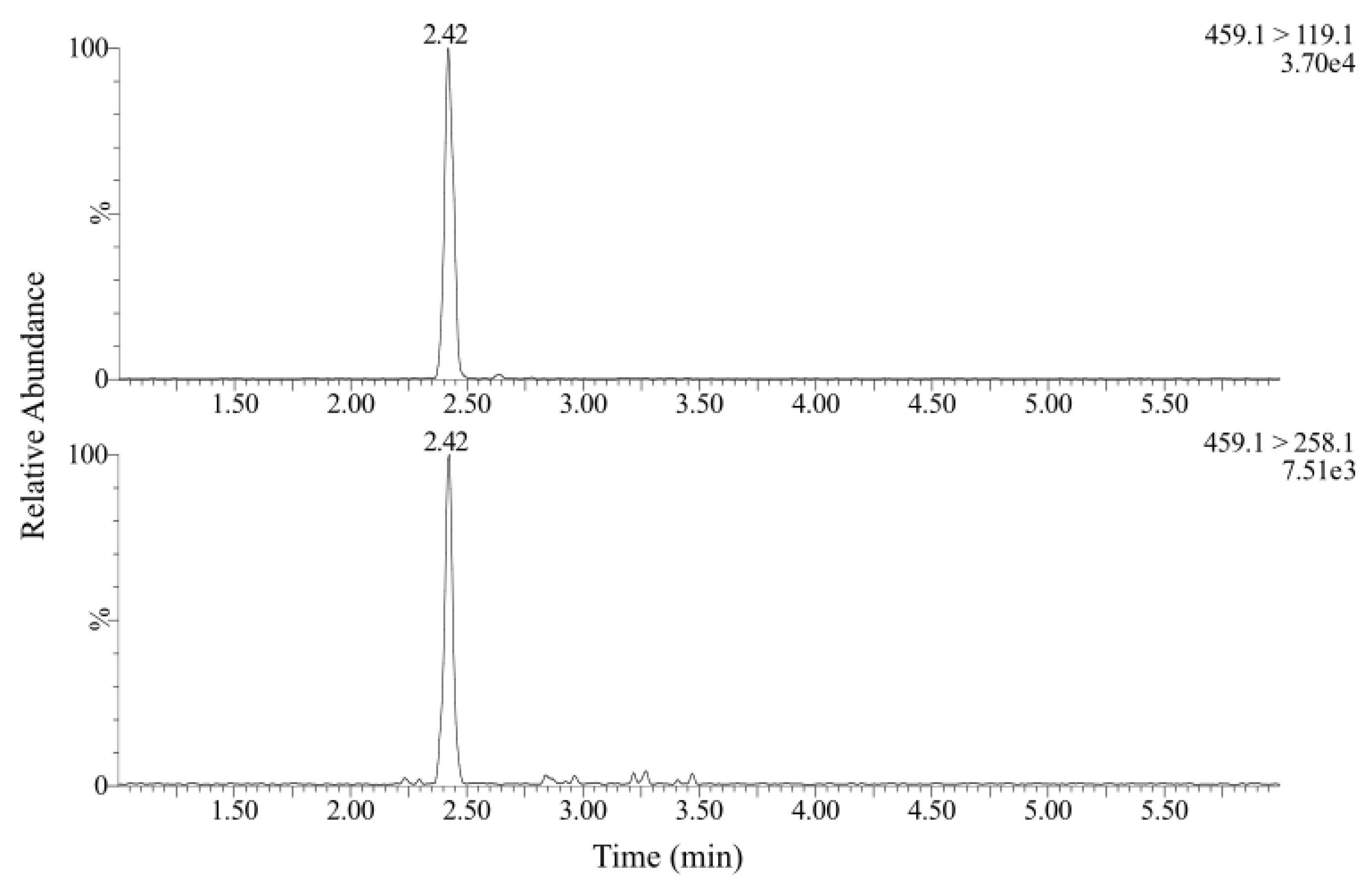
2. Materials and Methods
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Nunn, P.B. 50 years of research on α-amino-β-methylaminopropionic acid (β-methylaminoalanine). Phytochemistry 2017, 144, 271–281. [Google Scholar] [CrossRef]
- Spencer, P.S.; Nunn, P.B.; Hugon, J.; Ludolph, A.C.; Ross, S.M.; Roy, D.N.; Robertson, R.C. Guam Amyotrophic Lateral Sclerosis-Parkinsonism-Dementia linked to a plant excitant neurotoxin. Science 1987, 237, 517–522. [Google Scholar] [CrossRef]
- Cox, P.A.; Sacks, O.W. Cycad neurotoxins, consumption of flying foxes, and ALS-PDC disease in Guam. Neurology 2002, 58, 956–959. [Google Scholar] [CrossRef]
- Bradley, W.G.; Mash, D.C. Beyond Guam: The cyanobacteria/BMAA hypothesis of the cause of ALS and other neurodegenerative diseases. Amyotroph. Lateral Scler. 2009, 10, 7–20. [Google Scholar] [CrossRef]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Dietary exposure to an environmental toxin triggers neurofibrillary tangles and amyloid deposits in the brain. Proc. R. Soc. B Biol. Sci. 2016, 283, 20152397. [Google Scholar] [CrossRef] [Green Version]
- Cox, P.A.; Davis, D.A.; Mash, D.C.; Metcalf, J.S.; Banack, S.A. Do vervets and macaques respond differently to BMAA? Neurotoxicology 2016, 57, 310–311. [Google Scholar] [CrossRef]
- Lobner, D.; Piana, P.M.T.; Salous, A.K.; Peoples, R.W. β-N-methylamino-l-alanine enhances neurotoxicity through multiple mechanisms. Neurobiol. Dis. 2007, 25, 360–366. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Garamszegi, S.P.; Banack, S.A.; Dooley, P.D.; Coyne, T.M.; McLean, D.W.; Rotstein, D.S.; Mash, D.C.; Cox, P.A. BMAA, Methylmercury, and mechanisms of neurodegeneration in dolphins: A natural model of toxin exposure. Toxins 2021, 13, 697. [Google Scholar] [CrossRef]
- Cox, P.A.; Banack, S.A.; Murch, S.J.; Rasmussen, U.; Tien, G.; Bidigare, R.R.; Metcalf, J.S.; Morrison, L.F.; Codd, G.A.; Bergman, B. Diverse taxa of cyanobacteria produce β-N-methylamino-L-alanine, a neurotoxic amino acid. Proc. Natl. Acad. Sci. USA 2005, 102, 5074–5078. [Google Scholar] [CrossRef] [Green Version]
- Faassen, E.J.; Gillissen, F.; Lürling, M. A comparative study on three analytical methods for the determination of the neurotoxin BMAA in cyanobacteria. PLoS ONE 2012, 7, e36667. [Google Scholar] [CrossRef]
- Faassen, E.J. Presence of the neurotoxin BMAA in aquatic ecosystems: What do we really know? Toxins 2014, 6, 1109–1138. [Google Scholar] [CrossRef] [Green Version]
- Krüger, T.; Mönch, B.; Oppenhäuser, S.; Luckas, B. LC–MS/MS determination of the isomeric neurotoxins BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in cyanobacteria and seeds of Cycas revoluta and Lathyrus latifolius. Toxicon 2010, 55, 547–557. [Google Scholar] [CrossRef]
- Jiang, L.; Aigret, B.; De Borggraeve, W.M.; Spacil, Z.; Ilag, L.L. Selective LC-MS/MS method for the identification of BMAA from its isomers in biological samples. Anal. Bioanal. Chem. 2012, 403, 1719–1730. [Google Scholar] [CrossRef]
- Chernoff, N.; Hill, D.J.; Diggs, D.L.; Faison, B.D.; Francis, B.M.; Lang, J.R.; Larue, M.M.; Le, T.-T.; Loftin, K.A.; Lugo, J.N.; et al. A critical review of the postulated role of the non-essential amino acid, β-N-methylamino-L-alanine, in neurodegenerative disease in humans. J. Toxicol. Environ. Health Part B 2017, 20, 183–229. [Google Scholar] [CrossRef]
- Dunlop, R.A.; Banack, S.A.; Bishop, S.; Metcalf, J.S.; Murch, S.J.; Davis, D.A.; Stommel, E.W.; Karlsson, O.; Brittebo, E.B.; Chatziefthimiou, A.D.; et al. Is exposure to BMAA a risk factor for neurodegenerative diseases? A response to a critical review of the BMAA hypothesis. Neurotox. Res. 2021, 39, 81–106. [Google Scholar] [CrossRef]
- Jiang, L.; Eriksson, J.; Lage, S.; Jonasson, S.; Shams, S.; Mehine, M.; Ilag, L.L.; Rasmussen, U. Diatoms: A novel source for the neurotoxin BMAA in aquatic environments. PLoS ONE 2014, 9, e84578. [Google Scholar] [CrossRef]
- Wang, C.; Yan, C.; Qiu, J.; Liu, C.; Yan, Y.; Ji, Y.; Wang, G.; Chen, H.; Li, Y.; Li, A. Food web biomagnification of the neurotoxin β-N-methylamino-L-alanine in a diatom-dominated marine ecosystem in China. J. Hazard. Mater. 2021, 404, 124217. [Google Scholar] [CrossRef]
- Violi, J.P.; Facey, J.A.; Mitrovic, S.M.; Colville, A.; Rodgers, K.J. Production of β-methylamino-L-alanine (BMAA) and its isomers by freshwater diatoms. Toxins 2019, 11, 512. [Google Scholar] [CrossRef] [Green Version]
- Prego, R.; Guzmán-Zuñiga, D.; Varela, M.; de Castro, M.; Gómez-Gesteira, M. Consequences of winter upwelling events on biogeochemical and phytoplankton patterns in a western Galician ria (NW Iberian peninsula). Estuar. Coast. Shelf Sci. 2007, 73, 409–422. [Google Scholar] [CrossRef]
- Estrada, M. Phytoplankton distribution and composition off the coast of Galicia (northwest of Spain). J. Plankton Res. 1984, 6, 417–434. [Google Scholar] [CrossRef]
- Varela, M.; Prego, R.; Pazos, Y.; Moroño, Á. Influence of upwelling and river runoff interaction on phytoplankton assemblages in a Middle Galician Ria and comparison with northern and southern rias (NW Iberian Peninsula). Estuar. Coast. Shelf Sci. 2005, 64, 721–737. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Production of BMAA and DAB by diatoms (Phaeodactylum tricornutum, Chaetoceros sp., Chaetoceros calcitrans and, Thalassiosira pseudonana) and bacteria isolated from a diatom culture. Harmful Algae 2016, 58, 45–50. [Google Scholar] [CrossRef]
- Tilstone, G.H.; Miguez, B.M.; Figueiras, F.G. Diatom dynamics in a coastal ecosystem affected by upwelling: Coupling between species succession, circulation and biogeochemical processes. Mar. Ecol. Prog. Ser. 2000, 205, 23–41. [Google Scholar] [CrossRef] [Green Version]
- Christensen, S.J.; Hemscheidt, T.K.; Trapido-Rosenthal, H.; Laws, E.A.; Bidigare, R.R. Detection and quantification of β-methylamino-L-alanine in aquatic invertebrates. Limnol. Oceanogr. Methods 2012, 10, 891–898. [Google Scholar] [CrossRef]
- Jonasson, S.; Eriksson, J.; Berntzon, L.; Spáčil, Z.; Ilag, L.L.; Ronnevi, L.-O.; Rasmussen, U.; Bergman, B. Transfer of a cyanobacterial neurotoxin within a temperate aquatic ecosystem suggests pathways for human exposure. Proc. Natl. Acad. Sci. USA 2010, 107, 9252–9257. [Google Scholar] [CrossRef] [Green Version]
- Zguna, N.; Karlson, A.M.L.; Ilag, L.L.; Garbaras, A.; Gorokhova, E. Insufficient evidence for BMAA transfer in the pelagic and benthic food webs in the Baltic Sea. Sci. Rep. 2019, 9, 10406. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Kiselova, N.; Rosén, J.; Ilag, L.L. Quantification of neurotoxin BMAA (β-N-methylamino-L-alanine) in seafood from Swedish markets. Sci. Rep. 2014, 4, 6931. [Google Scholar] [CrossRef] [Green Version]
- Salomonsson, M.L.; Fredriksson, E.; Alfjorden, A.; Hedeland, M.; Bondesson, U. Seafood sold in Sweden contains BMAA: A study of free and total concentrations with UHPLC–MS/MS and dansyl chloride derivatization. Toxicol. Rep. 2015, 2, 1473–1481. [Google Scholar] [CrossRef] [Green Version]
- Braga, A.C.; Lage, S.; Pacheco, M.; Rydberg, S.; Costa, P.R. Native (Ruditapes decussatus) and non-indigenous (R. philippinarum) shellfish species living in sympatry: Comparison of regulated and non-regulated biotoxins accumulation. Mar. Environ. Res. 2017, 129, 147–155. [Google Scholar] [CrossRef]
- Lage, S.; Costa, P.R.; Moita, T.; Eriksson, J.; Rasmussen, U.; Rydberg, S.J. BMAA in shellfish from two Portuguese transitional water bodies suggests the marine dinoflagellate Gymnodinium catenatum as a potential BMAA source. Aquat. Toxicol. 2014, 152, 131–138. [Google Scholar] [CrossRef]
- Réveillon, D.; Séchet, V.; Hess, P.; Amzil, Z. Systematic detection of BMAA (β-N-methylamino-l-alanine) and DAB (2,4-diaminobutyric acid) in mollusks collected in shellfish production areas along the French coasts. Toxicon 2016, 110, 35–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondo, K.; Hammerschlag, N.; Basile, M.; Pablo, J.; Banack, S.A.; Mash, D.C. Cyanobacterial neurotoxin β-N-methylamino-L-alanine (BMAA) in shark fins. Mar. Drugs 2012, 10, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, M.; Karlsson, O.; Brandt, I. The environmental neurotoxin β-N-methylamino-l-alanine (l-BMAA) is deposited into birds’ eggs. Ecotoxicol. Environ. Saf. 2018, 147, 720–724. [Google Scholar] [CrossRef] [PubMed]
- Field, N.C.; Metcalf, J.S.; Caller, T.A.; Banack, S.A.; Cox, P.A.; Stommel, E.W. Linking β-methylamino-l-alanine exposure to sporadic amyotrophic lateral sclerosis in Annapolis, MD. Toxicon 2013, 70, 179–183. [Google Scholar] [CrossRef]
- Li, B.; Yu, S.; Li, G.; Chen, X.; Huang, M.; Liao, X.; Li, H.; Hu, F.; Wu, J. Transfer of a cyanobacterial neurotoxin, β-methylamino-l-alanine from soil to crop and its bioaccumulation in Chinese cabbage. Chemosphere 2019, 219, 997–1001. [Google Scholar] [CrossRef]
- Metcalf, J.S.; Banack, S.A.; Kotut, K.; Krienitz, L.; Codd, G.A. Amino acid neurotoxins in feathers of the Lesser Flamingo, Phoeniconaias minor. Chemosphere 2013, 90, 835–839. [Google Scholar] [CrossRef]
- Lance, E.; Arnich, N.; Maignien, T.; Biré, R. Occurrence of β-N-methylamino-l-alanine (BMAA) and isomers in aquatic environments and aquatic food sources for humans. Toxins 2018, 10, 83. [Google Scholar] [CrossRef] [Green Version]
- Dunlop, R.A.; Cox, P.A.; Banack, S.A.; Rodgers, K.J. The non-protein amino acid BMAA is misincorporated into human proteins in place of l-Serine causing protein misfolding and aggregation. PLoS ONE 2013, 8, e75376. [Google Scholar] [CrossRef] [Green Version]
- Xie, X.; Basile, M.; Mash, D.C. Cerebral uptake and protein incorporation of cyanobacterial toxin β-N-methylamino-L-alanine. Neuroreport 2013, 24, 779–784. [Google Scholar] [CrossRef]
- Glover, W.B.; Mash, D.C.; Murch, S.J. The natural non-protein amino acid N-β-methylamino-l-alanine (BMAA) is incorporated into protein during synthesis. Amino Acids 2014, 46, 2553–2559. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Powell, C.L.; Busman, M.; Doucette, G.J.; Moeller, P.D.R.; Silver, J.B.; Miller, P.E.; Hughes, M.P.; Singaram, S.; Silver, M.W.; et al. Detection of domoic acid in northern anchovies and California sea lions associated with an unusual mortality event. Nat. Toxins 1999, 7, 85–92. [Google Scholar] [CrossRef]
- Scholin, C.A.; Gulland, F.; Doucette, G.J.; Benson, S.; Busman, M.; Chavez, F.P.; Cordaro, J.; DeLong, R.; De Vogelaere, A.; Harvey, J.; et al. Mortality of sea lions along the central California coast linked to a toxic diatom bloom. Nature 2000, 403, 80–84. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Bargu, S.; Kieckhefer, T.; Silver, M.W. From sanddabs to blue whales: The pervasiveness of domoic acid. Toxicon 2002, 40, 971–977. [Google Scholar] [CrossRef]
- Lefebvre, K.A.; Quakenbush, L.; Frame, E.; Huntington, K.B.; Sheffield, G.; Stimmelmayr, R.; Bryan, A.; Kendrick, P.; Ziel, H.; Goldstein, T.; et al. Prevalence of algal toxins in Alaskan marine mammals foraging in a changing arctic and subarctic environment. Harmful Algae 2016, 55, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Davis, D.A.; Mondo, K.; Stern, E.; Annor, A.K.; Murch, S.J.; Coyne, T.M.; Brand, L.E.; Niemeyer, M.E.; Sharp, S.; Bradley, W.G.; et al. Cyanobacterial neurotoxin BMAA and brain pathology in stranded dolphins. PLoS ONE 2019, 14, e0213346. [Google Scholar] [CrossRef] [PubMed]
- López, A. Mammalia. In Inventario de la Biodiversidad Marina de Galicia: Proyecto LEMGAL; Bañón, R., Ed.; Consellería do Mar, Xunta de Galicia, Santiago de Compostela: Santiago de Compostela, Spain, 2017; pp. 567–570. [Google Scholar]
- Covelo, P.; López, A. First record of dwarf sperm whale (Kogia sima) in the north of Spain. Galemys Span. J. Mammal. 2021, 33, 64–69. [Google Scholar] [CrossRef]
- López, A.; Pierce, G.J.; Valeiras, X.; Santos, M.B.; Guerra, A. Distribution patterns of small cetaceans in Galician waters. J. Mar. Biol. Assoc. UK 2004, 84, 283–294. [Google Scholar] [CrossRef] [Green Version]
- Pierce, G.J.; Caldas, M.; Cedeira, J.; Santos, M.B.; Llavona, Á.; Covelo, P.; Martinez, G.; Torres, J.; Sacau, M.; López, A. Trends in cetacean sightings along the Galician coast, north-west Spain, 2003–2007, and inferences about cetacean habitat preferences. J. Mar. Biol. Assoc. UK 2010, 90, 1547–1560. [Google Scholar] [CrossRef]
- López, A.; Santos, M.B.; Pierce, G.J.; González, A.F.; Valeiras, X.; Guerra, A. Trends in strandings and by-catch of marine mammals in north-west Spain during the 1990s. J. Mar. Biol. Assoc. UK 2002, 82, 513–521. [Google Scholar] [CrossRef] [Green Version]
- Fontaine, M.C.; Roland, K.; Calves, I.; Austerlitz, F.; Palstra, F.P.; Tolley, K.A.; Ryan, S.; Ferreira, M.; Jauniaux, T.; Llavona, A.; et al. Postglacial climate changes and rise of three ecotypes of harbour porpoises, Phocoena phocoena, in western Palearctic waters. Mol. Ecol. 2014, 23, 3306–3321. [Google Scholar] [CrossRef]
- Kuiken, T.; Hartmann, M.G. European Cetacean Society: Pacific Grove. In Proceedings of the First ECS Workshop on Cetacean Pathology: Dissection Techniques and Tissue Sampling, Leiden, The Netherlands, 13–14 September 1991. [Google Scholar]
- Geraci, J.R.; Lounsbury, V.J. Marine Mammals Ashore: A Field Guide to Strandings: Texas A&M University Sea Grant Program; Publication TAMU-SG-93-601; Wildlife Disease Association: Lawrence, KS, USA, 1993. [Google Scholar]
- Vázquez, J.A.; De la Fuente, J.; Martínez-Cedeira, J.A.; Fernández, C.; Gozalbes, P.; López, A.; Arbelo, M. Documento Técnico Sobre Protocolo Nacional de Actuación para Varamientos de Cetáceos, Informe Realizado para el Ministerio de Agricultura, Alimentación y Medio Ambiente. 2015.
- Murch, S.J.; Cox, P.A.; Banack, S.A.; Steele, J.C.; Sacks, O.W. Occurrence of β-methylamino-l-alanine (BMAA) in ALS/PDC patients from Guam. Acta Neurol. Scand. 2004, 110, 267–269. [Google Scholar] [CrossRef] [PubMed]
- Masseret, E.; Banack, S.; Boumédiène, F.; Abadie, E.; Brient, L.; Pernet, F.; Juntas-Morales, R.; Pageot, N.; Metcalf, J.; Cox, P.; et al. Dietary BMAA exposure in an Amyotrophic Lateral Sclerosis Cluster from southern France. PLoS ONE 2013, 8, e83406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lage, S.; Burian, A.; Rasmussen, U.; Costa, P.R.; Annadotter, H.; Godhe, A.; Rydberg, S. BMAA extraction of cyanobacteria samples: Which method to choose? Environ. Sci. Pollut. Res. 2016, 23, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Lage, S.; Ström, L.; Godhe, A.; Rydberg, S. Kinetics of β-N-methylamino-L-alanine (BMAA) and 2, 4-diaminobutyric acid (DAB) production by diatoms: The effect of nitrogen. Eur. J. Phycol. 2019, 54, 115–125. [Google Scholar] [CrossRef]
- Wenzl, T.; Haedrich, J.; Schaechtele, A.; Robouch, P.; Stroka, J. Guidance Document on the Estimation of LOD and LOQ for Measurements in the Field of Contaminants in Feed and Food; Publications Office of the European Union: Luxembourg, 2016; Volume 58. [Google Scholar] [CrossRef]
- Lage, S.; Annadotter, H.; Rasmussen, U.; Rydberg, S. Biotransfer of β-N-Methylamino-l-alanine (BMAA) in a eutrophicated freshwater lake. Mar. Drugs 2015, 13, 1185–1201. [Google Scholar] [CrossRef] [Green Version]
- Błaszczyk, A.; Siedlecka-Kroplewska, K.; Woźniak, M.; Mazur-Marzec, H. Presence of ß-N-methylamino-L-alanine in cyanobacteria and aquatic organisms from waters of Northern Poland; BMAA toxicity studies. Toxicon 2021, 194, 90–97. [Google Scholar] [CrossRef]
- Karlsson, O.; Berg, C.; Brittebo, E.B.; Lindquist, N.G. Retention of the cyanobacterial neurotoxin β-N-methylamino-l-alanine in melanin and neuromelanin-containing cells—A possible link between Parkinson-dementia complex and pigmentary retinopathy. Pigment Cell Melanoma Res. 2009, 22, 120–130. [Google Scholar] [CrossRef]
- Masuoka, D.T.; Alcaraz, A.F.; Cohen, M.B.; Spolter, L. Acute distribution of 14C-amino acids in mice as determined by whole-body autoradiography: Adjunct for radio-pharmaceutical synthesis. Int. J. Appl. Radiat. Isot. 1973, 24, 705–706. [Google Scholar] [CrossRef]
- Karlsson, O.; Jiang, L.; Andersson, M.; Ilag, L.L.; Brittebo, E.B. Protein association of the neurotoxin and non-protein amino acid BMAA (β-N-methylamino-l-alanine) in the liver and brain following neonatal administration in rats. Toxicol. Lett. 2014, 226, 1–5. [Google Scholar] [CrossRef]
- Jiao, Y.; Chen, Q.; Chen, X.; Wang, X.; Liao, X.; Jiang, L.; Wu, J.; Yang, L. Occurrence and transfer of a cyanobacterial neurotoxin β-methylamino-l-alanine within the aquatic food webs of Gonghu Bay (Lake Taihu, China) to evaluate the potential human health risk. Sci. Total Environ. 2014, 468–469, 457–463. [Google Scholar] [CrossRef]
- Hammerschlag, N.; Davis, D.A.; Mondo, K.; Seely, M.S.; Murch, S.J.; Glover, W.B.; Divoll, T.; Evers, D.C.; Mash, D.C. Cyanobacterial neurotoxin BMAA and mercury in sharks. Toxins 2016, 8, 238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mondo, K.; Glover, W.B.; Murch, S.J.; Liu, G.; Cai, Y.; Davis, D.A.; Mash, D.C. Environmental neurotoxins β-N-methylamino-l-alanine (BMAA) and mercury in shark cartilage dietary supplements. Food Chem. Toxicol. 2014, 70, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Faassen, E.J.; Antoniou, M.G.; Beekman-Lukassen, W.; Blahova, L.; Chernova, E.; Christophoridis, C.; Combes, A.; Edwards, C.; Fastner, J.; Harmsen, J.; et al. A collaborative evaluation of LC-MS/MS based methods for BMAA analysis: Soluble bound BMAA found to be an important fraction. Mar. Drugs 2016, 14, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murch, S.J.; Cox, P.A.; Banack, S.A. A mechanism for slow release of biomagnified cyanobacterial neurotoxins and neurodegenerative disease in Guam. Proc. Natl. Acad. Sci. USA 2004, 101, 12228–12231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.; Xue, J.; Park, K.J.; Kannan, K.; Moon, H.-B. Tissue-specific accumulation and body burden of parabens and their metabolites in small cetaceans. Environ. Sci. Technol. 2019, 53, 475–481. [Google Scholar] [CrossRef]
- Tilstone, G.H.; Figueiras, F.G.; Lorenzo, L.M. Phytoplankton composition, photosynthesis and primary production during different hydrographic conditions at the Northwest Iberian upwelling system. Mar. Ecol. Prog. Ser. 2003, 252, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Lorenzo, L.M.; Arbones, B.; Tilstone, G.H.; Figueiras, F.G. Across-shelf variability of phytoplankton composition, photosynthetic parameters and primary production in the NW Iberian upwelling system. J. Mar. Syst. 2005, 54, 157–173. [Google Scholar] [CrossRef]
- Villamaña, M.; Marañón, E.; Cermeño, P.; Estrada, M.; Fernández-Castro, B.; Figueiras, F.G.; Latasa, M.; Otero-Ferrer, J.L.; Reguera, B.; Mouriño-Carballido, B. The role of mixing in controlling resource availability and phytoplankton community composition. Prog. Oceanogr. 2019, 178, 102181. [Google Scholar] [CrossRef]
- Gallego, J.R.; González-Rojas, E.; Peláez, A.I.; Sánchez, J.; García-Martínez, M.J.; Ortiz, J.E.; Torres, T.; Llamas, J.F. Natural attenuation and bioremediation of Prestige fuel oil along the Atlantic coast of Galicia (Spain). Org. Geochem. 2006, 37, 1869–1884. [Google Scholar] [CrossRef]
- Calvo, S.M.; Dosil, J.; del Carmen López Rodríguez, M.; Criado, I.B.; Cremades, J. Checklist of the benthic marine and brackish Galician algae (NW Spain). An. Jardín Botánico Madr. 2005, 62, 69–100. [Google Scholar] [CrossRef]
- European Union Commission Regulation(EC) No 853/2004 of the European Parliament and of the Council of 29 April 2004 Laying Down Specific Hygiene Rules for on the Hygiene of Foodstuffs 2004. Available online: https://www.legislation.gov.uk/eur/2004/853/contents (accessed on 23 December 2021).



{kind=link}
{kind=link}
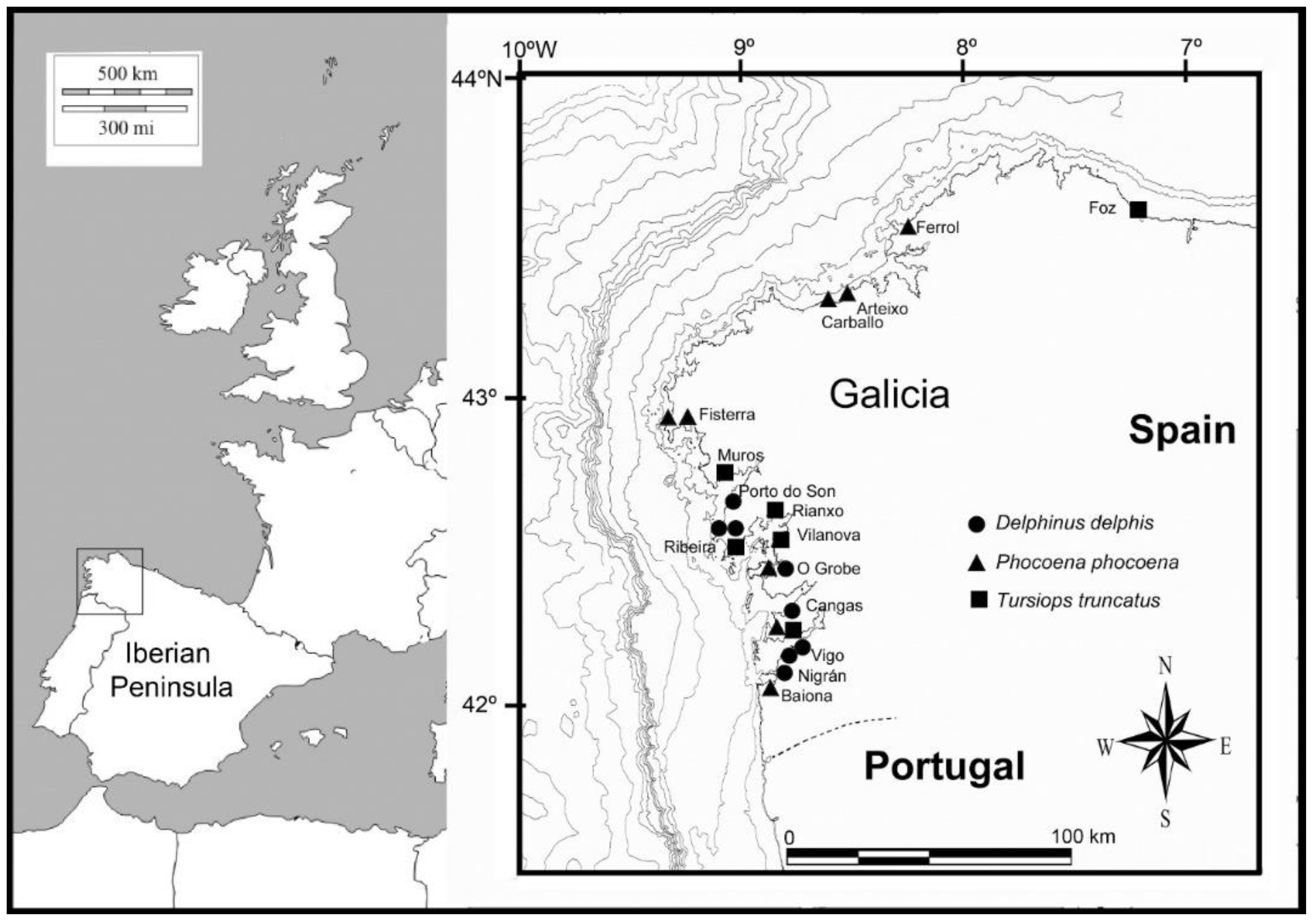
| Sample ID | Species | Data of Collection (yyyy/mm/dd) | Location (Locality and Coordinates) | Size (cm) * | Female (F)/Male (M) | Degradation State | Observations |
|---|---|---|---|---|---|---|---|
| DDE194 | D. delphis | 2017/07/16 | Porto do Son (42.68131, −9.03000) | 194 | F | 3 | No signs of bycatch |
| DDE195 | D. delphis | 2017/07/16 | Ribeira (42.56142222, −8.987644444) | 195 | M | 3 | No signs of bycatch |
| DDE189 | D. delphis | 2017/07/31 | Cangas (42.24911, −8.79091) | 189 | F | 3 | Signs of bycatch |
| DDE182 | D. delphis | 2017/07/31 | Nigrán (42.14245, −8.83796) | 182 | M | 3 | Three broken ribs and subepidermic hematoma |
| DDE173 | D. delphis | 2018/06/09 | Ribeira (42.57156, −9.07536) | 173 | M | 1 | No signs of bycatch |
| DDE156 | D. delphis | 2018/11/07 | Vigo (42.19025, −8.80904) | 156 | M | 2 | - |
| DDE124 | D. delphis | 2018/11/25 | O Grove (42.45576667, −8.921633333) | 124 | F | 3 | Signs of bycatch |
| DDE172 | D. delphis | 2018/12/17 | Vigo (42.22298056, −8.772766667) | 172 | F | 3 | Signs of bycatch |
| PPH153 | P. phocoena | 2009/11/25 | Fisterra (42.941181, −9.231806) | 153 | F | 3 | Good aspect |
| PPH127 | P. phocoena | 2011/02/07 | Fisterra (42.908431, −9.258883) | 127 | M | 3 | Signs of bycatch |
| PPH104 | P. phocoena | 2011/05/27 | Baiona (43.056527, −9.295471) | 104 | F | 2 | Signs of bycatch |
| PPH137 | P. phocoena | 2012/12/02 | Arteixo (43.316153, −8.534233) | 137 | F | 3 | Signs gulls and shark bites |
| PPH142 | P. phocoena | 2013/10/15 | O Grove (43.463064, −8.332825) | 142 | F | 3 | - |
| PPH159 | P. phocoena | 2014/02/28 | Ferrol (43.540556, −8.298353) | 159 | M | 3 | Signs of bycatch |
| PPH131 | P. phocoena | 2015/01/16 | Cangas (42.261026, −8.849869) | 131 | M | 3 | Signs of bycatch |
| PPH160 | P. phocoena | 2017/02/20 | Carballo (43.456950, −8.673272) | 160 | M | 3 | Signs of bycatch |
| TTR258 | T. truncatus | 2015/10/30 | Ribeira (42.523304, −9.014026) | 258 | M | 3 | Stomach with food and Anisakis, ulcers |
| TTR314 | T. struncatus | 2015/12/30 | Vilanova de Arousa (42.570726, −8.830053) | 314 | F | 2 | Empty stomach |
| TTR286 | T. truncatus | 2016/09/02 | Cangas (42.295522, −8.820978 | 286 | F | 3 | Pregnant |
| TTR150.5 | T. truncatus | 2016/09/11 | Rianxo (42.64833056, −8.8258) | 150.5 | M | 3 | Signs of aggressions (possible infanticide) |
| TTR275 | T. truncatus | 2017/06/19 | Foz (43.5665472, −7.2546361) | 275 | F | 2 | Skinny |
| TTR138 | T. truncatus | 2018/11/30 | Muros (42.7747527, −9.05476) | −138 | M | 4 | Signs of bycatch |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soliño, L.; Kim, S.-Y.; López, A.; Covelo, P.; Rydberg, S.; Costa, P.R.; Lage, S. No β-N-Methylamino-L-alanine (BMAA) Was Detected in Stranded Cetaceans from Galicia (North-West Spain). J. Mar. Sci. Eng. 2022, 10, 314. https://doi.org/10.3390/jmse10030314
Soliño L, Kim S-Y, López A, Covelo P, Rydberg S, Costa PR, Lage S. No β-N-Methylamino-L-alanine (BMAA) Was Detected in Stranded Cetaceans from Galicia (North-West Spain). Journal of Marine Science and Engineering. 2022; 10(3):314. https://doi.org/10.3390/jmse10030314
Chicago/Turabian StyleSoliño, Lucía, Sea-Yong Kim, Alfredo López, Pablo Covelo, Sara Rydberg, Pedro Reis Costa, and Sandra Lage. 2022. "No β-N-Methylamino-L-alanine (BMAA) Was Detected in Stranded Cetaceans from Galicia (North-West Spain)" Journal of Marine Science and Engineering 10, no. 3: 314. https://doi.org/10.3390/jmse10030314