
Seasonal Controls of Seawater CO2 Systems in Subtropical Coral Reefs: A Case Study from the Eastern Coast of Shenzhen, China
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Sampling and Analyses
2.3. Incubation Experiments of Coral Metabolism
2.4. Estimation of CO2 Fluxes
2.5. Budget Model to Distinguish Controlling Processes of CO2 System
2.6. Statistical Analysis
3. Results
3.1. Hydrological Features
3.2. Spatiotemporal Variations in Marine CO2 System
3.2.1. TA and DIC
3.2.2. pHT@in situ and ΩA@in situ
3.2.3. pCO2@in situ and FCO2
3.3. Coral Metabolic Process
4. Discussion
4.1. Comparison of CO2 Parameters with Other Reefs around the World
4.2. Factors Controlling the Variability of Seawater CO2 Parameters
4.2.1. Temperature
4.2.2. Water Mixing
4.2.3. Photosynthesis and Respiration
4.2.4. Coral Metabolic Activity
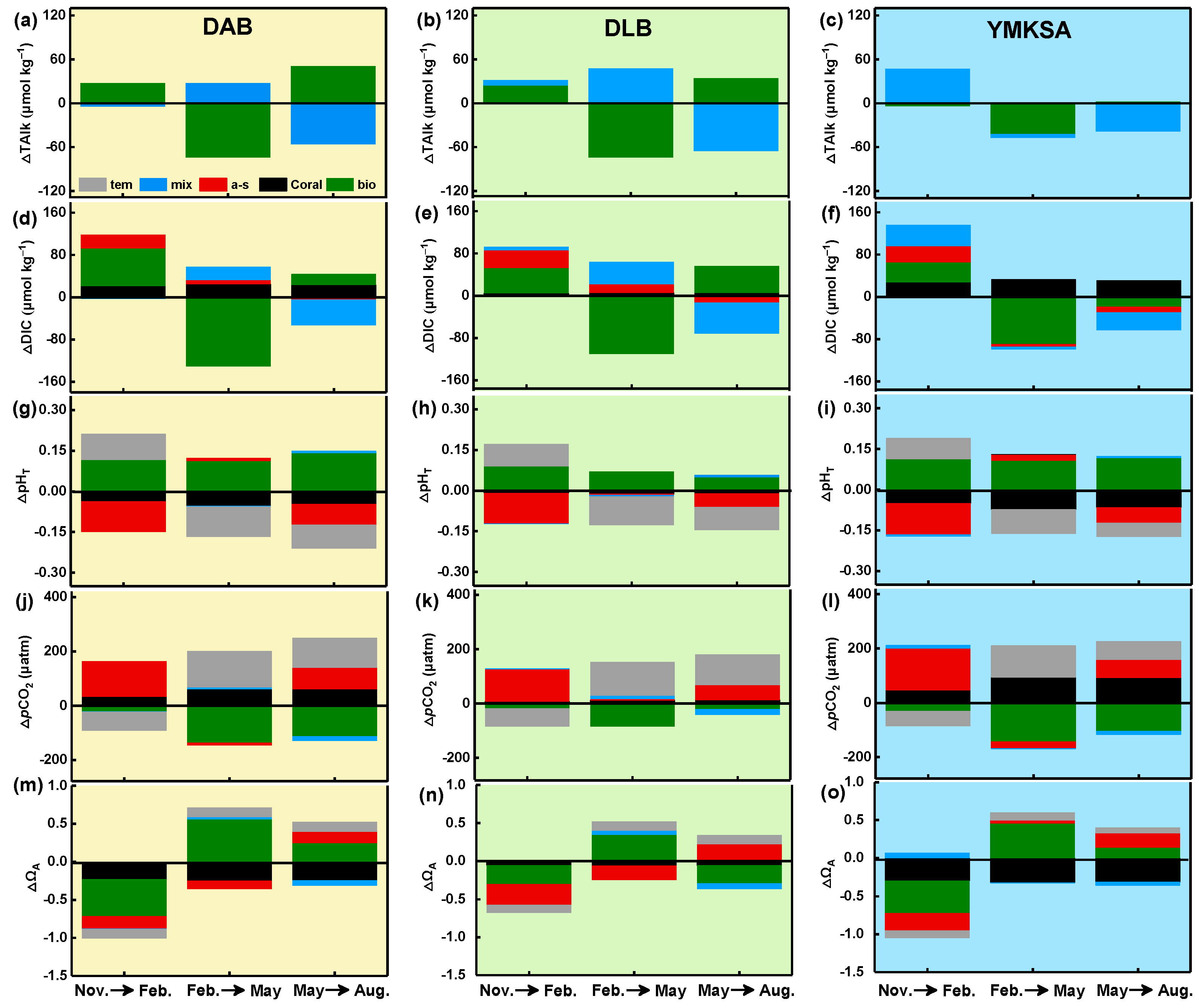
4.3. 1–D Model Reveals the Seasonal Driving Mechanism of CO2 System
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NOAA. Trends in Atmosphere Carbon Dioxide, Global Greenhouse Gas Reference Network. 2023. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/global.html (accessed on 28 May 2023).
- Feely, R.A.; Sabine, C.L.; Lee, K.; Berelson, W.; Kleypas, J.; Fabry, V.; Millero, F.J. Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 2004, 305, 362–363. [Google Scholar] [CrossRef] [PubMed]
- Doney, S.C.; Fabry, V.J.; Feely, R.A.; Kleypas, J.A. Ocean acidification: The other CO2 problem. Annu. Rev. Mar. Sci. 2009, 1, 169–192. [Google Scholar] [CrossRef] [PubMed]
- Riebesell, U.; Kortzinger, A.; Oschlies, A. Sensitivities of marine carbon fluxes to ocean change. Proc. Natl. Acad. Sci. USA 2009, 106, 20602–20609. [Google Scholar] [CrossRef]
- Inoue, S.; Kayanne, H.; Yamamoto, S.; Kurihara, H. Spatial community shift from hard to soft corals in acidified water. Nat. Clim. Change 2013, 3, 683–687. [Google Scholar] [CrossRef]
- Enochs, I.C.; Manzello, D.P.; Donham, E.M.; Kolodziej, G.; Okano, R.; Johnston, L.; Price, N.N. Shift from coral to macroalgae dominance on a volcanically acidified reef. Nat. Clim. Change 2015, 5, 1083–1088. [Google Scholar] [CrossRef]
- Horwitz, R.; Hoogenboom, M.O.; Fine, M. Spatial competition dynamics between reef corals under ocean acidification. Sci. Rep. 2017, 7, 40288. [Google Scholar] [CrossRef]
- Agostini, S.; Harvey, B.P.; Wada, S.; Kon, K.; Milazzo, M.; Inaba, K.; Hall-Spencer, J.M. Ocean acidification drives community shifts towards simplified non-calcified habitats in a subtropical-temperate transition zone. Sci. Rep. 2018, 8, 11354. [Google Scholar] [CrossRef]
- Page, H.N.; Courtney, T.A.; De Carlo, E.H.; Howins, N.M.; Koester, I.; Andersson, A.J. Spatiotemporal variability in seawater carbon chemistry for a coral reef flat in Kāne‘ohe Bay, Hawai‘i. Limnol. Oceanogr. 2019, 64, 913–934. [Google Scholar] [CrossRef]
- Kurihara, H.; Watanabe, A.; Tsugi, A.; Mimura, I.; Hongo, C.; Kawai, T.; Golbuu, Y. Potential local adaptation of corals at acidified and warmed Nikko Bay, Palau. Sci. Rep. 2021, 11, 11192. [Google Scholar] [CrossRef]
- Orr, J.C.; Fabry, V.J.; Aumont, O.; Bopp, L.; Doney, S.C.; Feely, R.A.; Yool, A. Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 2005, 437, 681–686. [Google Scholar] [CrossRef]
- Bates, N.R.; Astor, Y.M.; Church, M.J.; Currie, K.; Dore, J.E.; González-Dávila, M.; Santana-Casiano, J.M. A time-series view of changing surface ocean chemistry due to ocean uptake of anthropogenic CO2 and ocean acidification. Oceanography 2014, 27, 126–141. [Google Scholar] [CrossRef]
- Lauvset, S.K.; Gruber, N.; Landschützer, P.; Olsen, A.; Tjiputra, J. Trends and drivers in global surface ocean pH over the past 3 decades. Biogeosciences 2015, 12, 1285–1298. [Google Scholar] [CrossRef]
- Dumousseaud, C.; Achterberg, E.P.; Tyrrell, T.; Charalampopoulou, A.; Schuster, U.; Hartman, M.; Hydes, D.J. Contrasting effects of temperature and winter mixing on the seasonal and inter-annual variability of the carbonate system in the Northeast Atlantic Ocean. Biogeosciences 2010, 7, 1481–1492. [Google Scholar] [CrossRef]
- Artioli, Y.; Blackford, J.C.; Butenschön, M.; Holt, J.T.; Wakelin, S.L.; Thomas, H.; Allen, J.I. The carbonate system in the North Sea: Sensitivity and model validation. J. Mar. Sys. 2012, 102, 1–13. [Google Scholar] [CrossRef]
- Wimart-Rousseau, C.; Lajaunie-Salla, K.; Marrec, P.; Wagener, T.; Raimbault, P.; Lagadec, V.; Lefèvre, D. Temporal variability of the carbonate system and air-sea CO2 exchanges in a Mediterranean human-impacted coastal site. Estuar. Coast. Shelf Sci. 2020, 236, 106641. [Google Scholar] [CrossRef]
- Ghosh, J.; Chakraborty, K.; Bhattacharya, T.; Valsala, V.; Baduru, B. Impact of coastal upwelling dynamics on the pCO2 variability in the southeastern Arabian Sea. Prog. Oceanogr. 2022, 203, 102785. [Google Scholar] [CrossRef]
- Takeshita, Y.; Frieder, C.A.; Martz, T.R.; Ballard, J.R.; Feely, R.A.; Kram, S. Including high-frequency variability in coastal ocean acidification projections. Biogeosciences 2015, 12, 5853–5870. [Google Scholar] [CrossRef]
- Li, C.L.; Zhai, W.D. Decomposing monthly declines in subsurface-water pH and aragonite saturation state from spring to autumn in the North Yellow Sea. Cont. Shelf Res. 2019, 185, 37–50. [Google Scholar] [CrossRef]
- Alin, S.R.; Allen, B.; Suhrbier, A.; Davis, J.; Feely, R.A.; Newton, J. Water chemistry, larval oysters, and ocean acidification in a complex, urbanized estuary (Puget Sound, Washington). J. Shellfish. Res. 2012, 31, 259. [Google Scholar]
- Wallace, R.B.; Baumann, H.; Grear, J.S.; Aller, R.C.; Gobler, C.J. Coastal ocean acidification: The other eutrophication problem. Estuar. Coast. Shelf Sci. 2014, 148, 1–13. [Google Scholar] [CrossRef]
- Ferrera, C.M.; Jacinto, G.S.; Chen, C.; Diego-Mcglone, M.; Datoc, M.; Lagumen, M. Carbonate parameters in high and low productivity areas of the Sulu Sea, Philippines. Mar. Chem. 2017, 195, 2–14. [Google Scholar] [CrossRef]
- Li, C.L.; Yang, D.Z.; Zhai, W.D. Effects of warming, eutrophication and climate variability on acidification of the seasonally stratified North Yellow Sea over the past 40 years. Sci. Total Environ. 2022, 815, 152935. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Liu, J.; Uthaipan, K.; Song, X.; Xu, Y.; He, B.; Dai, M. Dynamics of inorganic carbon and pH in a large subtropical continental shelf system: Interaction between eutrophication, hypoxia, and ocean acidification. Limnol. Oceanogr. 2020, 65, 1359–1379. [Google Scholar] [CrossRef]
- Cai, W.J.; Hu, X.; Huang, W.J.; Murrell, M.C.; Lehrter, J.C.; Lohrenz, S.E. Acidification of subsurface coastal waters enhanced by eutrophication. Nat. Geosci. 2011, 4, 766–770. [Google Scholar] [CrossRef]
- Yang, B.; Gao, X.; Zhao, J.; Liu, Y.; Lui, H.K.; Huang, T.H.; Chen, C.; Xing, Q.G. Massive shellfish farming might accelerate coastal acidification: A case study on carbonate system dynamics in a bay scallop (Argopecten irradians) farming area, North Yellow Sea. Sci. Total Environ. 2021, 798, 149214. [Google Scholar] [CrossRef]
- Manzello, D.P. Ocean acidification hotspots: Spatiotemporal dynamics of the seawater CO2 system of eastern Pacific coral reefs. Limnol. Oceanogr. 2010, 55, 239–248. [Google Scholar] [CrossRef]
- Yang, B.; Zhang, Z.; Cui, Z.; Chen, B.; Zheng, H.; Xiao, B. Multiple factors driving carbonate system in subtropical coral community environments along Dapeng Peninsula, South China Sea. Atmosphere 2023, 14, 688. [Google Scholar] [CrossRef]
- Allison, N. Reconstructing coral calcification fluid dissolved inorganic carbon chemistry from skeletal boron: An exploration of potential controls on coral aragonite B/Ca. Heliyon 2017, 3, e00387. [Google Scholar] [CrossRef] [PubMed]
- Cotovicz, L.C., Jr.; Chielle, R.; Marins, R.V. Air-sea CO2 flux in an equatorial continental shelf dominated by coral reefs (Southwestern Atlantic Ocean). Cont. Shelf Res. 2020, 204, 104175. [Google Scholar] [CrossRef]
- Dong, X.; Qi, D.; Chen, B.; Wu, Y.; Zheng, X.; Lin, H. Differential roles of anthropogenic CO2 in mediating seasonal amplitudes of ocean acidification metrics over a coastal coral habitat. J. Marine Syst. 2024, 241, 103910. [Google Scholar] [CrossRef]
- Zhang, C.; Huang, H.; Ye, C.; Huang, L.; Li, X.; Lian, J.; Liu, S. Diurnal and seasonal variations of carbonate system parameters on Luhuitou fringing reef, Sanya Bay, Hainan Island, South China Sea. Deep Sea Res. Pt. II 2013, 96, 65–74. [Google Scholar] [CrossRef]
- Chen, T.R.; Yu, K.F.; Shi, Q. Twenty-five years of change in scleractinian coral communities of Daya Bay (northern South China Sea) and its response to the 2008 AD extreme cold climate event. Chin. Sci. Bull. 2009, 54, 812–820. [Google Scholar] [CrossRef]
- Zhou, P.; Li, D.; Li, H.; Fang, H.; Huang, C.; Zhang, Y.; Yang, J. Distribution of radionuclides in a marine sediment core off the waterspout of the nuclear power plants in Daya Bay, northeastern South China Sea. J. Environ. Radioactiv. 2015, 145, 102–112. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Wang, L.; Wang, X.; Van Damme, K.; Ning, J.; Li, Y.; Du, F. Spatial and temporal variabilities of coastal nekton community structure and phylogenetic diversity in Daya and Dapeng Bay, southern China. Ecol. Indic. 2021, 131, 108226. [Google Scholar] [CrossRef]
- Dickson, A.G.; Sabine, C.L.; Christian, J.R. Guide to Best Practices for Ocean CO2 Measurements; North Pacific Marine Science Organization: Victoria, BC, Canada, 2007. [Google Scholar]
- Pelletier, G.J.; Lewis, E.; Wallace, D.W.R. CO2SYS.XLS: A Calculator for the CO2 System in Seawater for Microsoft Excel/VBA, version 24; Washington State Department of Ecology: Olympia, WA, USA, 2015.
- Meng, L.; Huang, W.; Yang, E.G. High temperature bleaching events can increase thermal tolerance of Porites lutea in the Weizhou Island. Haiyang Xuebao 2022, 44, 87–96. [Google Scholar]
- Wanninkhof, R. Relationship between wind speed and gas exchange over the ocean. J. Geophys. Res. Ocean. 1992, 97, 7373–7382. [Google Scholar] [CrossRef]
- Sweeney, C.; Gloor, E.; Jacobson, A.R.; Key, R.M.; McKinley, G.; Sarmiento, J.L. Constraining global air-sea gas exchange for CO2 with recent bomb 14C measurements. Glob. Biogeochem. Cycles 2007, 21, 1–10. [Google Scholar] [CrossRef]
- South China Sea and Adjacent Seas Data Center. 2022. Available online: http://ocean.geodata.cn (accessed on 28 May 2023).
- Xue, L.; Cai, W.J.; Hu, X.; Sabine, C.; Jones, S.; Sutton, A.J. Sea surface carbon dioxide at the Georgia time series site (2006–2007): Air-sea flux and controlling processes. Prog. Oceanogr. 2016, 140, 14–26. [Google Scholar] [CrossRef]
- Xue, L.; Cai, W.J.; Sutton, A.J.; Sabine, C. Sea surface aragonite saturation state variations and control mechanisms at the Gray’s Reef time-series site off Georgia, USA (2006–2007). Mar. Chem. 2017, 195, 27–40. [Google Scholar] [CrossRef]
- Dai, M.H.; Guo, X.; Zhai, W.; Yuan, L.Y.; Wang, B.; Tang, T.T. Oxygen depletion in the upper reach of the Pearl River estuary during a winter drought. Mar. Chem. 2006, 102, 159–169. [Google Scholar] [CrossRef]
- DeCarlo, T.M.; Cohen, A.L.; Wong, G.T.; Shiah, F.K.; Lentz, S.J.; Davis, K.A.; Lohmann, P. Community production modulates coral reef pH and the sensitivity of ecosystem calcification to ocean acidification. J. Geophys. Res.Ocean. 2017, 122, 745–761. [Google Scholar] [CrossRef]
- Guo, Y.J.; Zhou, W.H.; Yuan, X.C. Responses of two species of reef-building corals to acidification and dissolved organic carbon enrichment. J. Trop. Oceanogr. 2018, 7, 57–63. [Google Scholar]
- Kuffner, I.B.; Hickey, T.D.; Morrison, J.M. Calcification rates of the massive coral Siderastrea siderea and crustose coralline algae along the Florida Keys (USA) outer-reef tract. Coral Reefs 2013, 32, 987–997. [Google Scholar] [CrossRef]
- Nishida, K.; Ishikawa, K.; Iguchi, A.; Tanaka, Y.; Sato, M.; Ishimura, T.; Suzuki, A. Skeletal oxygen and carbon isotope compositions of Acropora coral primary polyps experimentally cultured at different temperatures. Geochem. Geophy. Geosy. 2014, 15, 2840–2849. [Google Scholar] [CrossRef]
- Chan, W.Y.; Eggins, S.M. Calcification responses to diurnal variation in seawater carbonate chemistry by the coral Acropora formosa. Coral Reefs 2017, 36, 763–772. [Google Scholar] [CrossRef]
- Zhao, H.; Zhang, J.L.; Wang, H. Preliminary study on carbon sequestration capacity of two hermatypic corals. J. Trop. Oceanogr. 2023. [Google Scholar] [CrossRef]
- Fagan, K.E.; Mackenzie, F.T. Air–sea CO2 exchange in a subtropical estuarine-coral reef system, Kaneohe Bay, Oahu, Hawaii. Mar. Chem. 2007, 106, 174–191. [Google Scholar] [CrossRef]
- Akhand, A.; Watanabe, K.; Chanda, A.; Tokoro, T.; Kuwae, T. Lateral carbon fluxes and CO2 evasion from a subtropical mangrove-seagrass-coral continuum. Sci. Total Environ. 2021, 752, 142190. [Google Scholar] [CrossRef]
- Zhang, Y.L.; Zou, R.L. Study on community structure of shallow water stony coral in Daya Bay. Tropic. Oceanol. 1987, 6, 13–18. [Google Scholar]
- Yan, H.; Yu, K.; Shi, Q.; Tan, Y.; Zhang, H.; Zhao, M.; Li, S.; Chen, T.; Huang, L.; Wang, P. Coral reef ecosystems in the South China Sea as a source of atmospheric CO2 in summer. Chin. Sci. Bull. 2011, 56, 676–684. [Google Scholar] [CrossRef]
- Lonborg, C.; Calleja, M.L.; Fabricius, K.E.; Smith, J.N.; Achterberg, E.P. The great barrier reef: A source of CO2 to the at-mosphere. Mar. Chem. 2019, 210, 24–33. [Google Scholar] [CrossRef]
- Hannan, K.D.; Miller, G.M.; Watson, S.A.; Rummer, J.L.; Fabricius, K.; Munday, P.L. Diel pCO2 variation among coral reefs and microhabitats at Lizard Island, Great Barrier Reef. Coral Reefs 2020, 39, 1391–1406. [Google Scholar] [CrossRef]
- Albright, R.; Langdon, C.; Anthony, K.R.N. Dynamics of seawater carbonate chemistry, production, and calcification of a coral reef flat, central Great Barrier Reef. Biogeosciences 2013, 10, 6747–6758. [Google Scholar] [CrossRef]
- Norzagaray, C.O.; Hernandez-Ayon, J.M.; Castro, R.; Calderon-Aguilera, L.E.; Martz, T.; Valdivieso-Ojeda, J.A.; Lara-Lara, R. Seasonal controls of the carbon biogeochemistry of a fringing coral reef in the Gulf of California, Mexico. Cont. Shelf Res. 2020, 211, 104279. [Google Scholar] [CrossRef]
- Knebel, O.; Carvajal, C.; Kench, P.; Gehrels, R. Spatial pH variability of coral reef flats of Kiritimati Island, Kiribati. Mar. Environ. Res. 2023, 185, 105861. [Google Scholar] [CrossRef]
- Weiss, R.F. Carbon dioxide in water and seawater: The solubility of a non-ideal gas. Mar. Chem. 1974, 2, 203–215. [Google Scholar] [CrossRef]
- Albuquerque, C.; Kerr, R.; Monteiro, T.; Orselli, I.B.M.; de Carvalho-Borges, M.; de Oliveira Carvalho, A.D.C.; Mendes, C.R.B. Seasonal variability of carbonate chemistry and its controls in a subtropical estuary. Estuar. Coast. Shelf Sci. 2022, 276, 108020. [Google Scholar] [CrossRef]
- Hu, X.; Nuttall, M.F.; Wang, H.; Yao, H.; Staryk, C.J.; McCutcheon, M.R. Seasonal variability of carbonate chemistry and decadal changes in waters of a marine sanctuary in the Northwestern Gulf of Mexico. Mar. Chem. 2018, 205, 16–28. [Google Scholar] [CrossRef]
- McCutcheon, M.R.; Yao, H.; Staryk, C.J.; Hu, X. Temporal variability and driving factors of the carbonate system in the Aransas Ship Channel, TX, USA: A time series study. Biogeosciences 2021, 18, 4571–4586. [Google Scholar] [CrossRef]
- Deng, X.; Zhang, G.L.; Xin, M.; Liu, C.Y.; Cai, W.J. Carbonate chemistry variability in the southern Yellow Sea and East China Sea during spring of 2017 and summer of 2018. Sci. Total Environ. 2021, 779, 146376. [Google Scholar] [CrossRef]
- Yang, W.; Guo, X.; Cao, Z.; Xu, Y.; Wang, L.; Guo, L.; Dai, M. Seasonal dynamics of the carbonate system under complex circulation schemes on a large continental shelf: The northern South China Sea. Prog. Oceanogr. 2021, 197, 102630. [Google Scholar] [CrossRef]
- Liang, B.; Xiu, P.; Hu, J.; Li, S. Seasonal and spatial controls on the eutrophication-induced acidification in the Pearl River Estuary. J. Geophys. Res. Ocean. 2021, 126, e2020JC017107. [Google Scholar] [CrossRef]
- Chou, W.C.; Tishchenko, P.Y.; Chuang, K.Y.; Gong, G.G.; Shkirnikova, E.M.; Tishchenko, P.P. The contrasting behaviors of CO2 systems in river-dominated and ocean-dominated continental shelves: A case study in the East China Sea and the Peter the Great Bay of Japan/East Sea in summer 2014. Mar. Chem. 2017, 195, 50–60. [Google Scholar] [CrossRef]
- Wright-Fairbanks, E.K.; Saba, G.K. Quantification of the Dominant Drivers of Acidification in the Coastal Mid-Atlantic Bight. J. Geophys. Res. Ocean. 2022, 127, e2022JC018833. [Google Scholar] [CrossRef]
- Henson, H.C.; Holding, J.M.; Meire, L.; Rysgaard, S.; Stedmon, C.A.; Stuart-Lee, A.; Sejr, M. Coastal freshening drives acidification state in Greenland fjords. Sci. Total Environ. 2023, 855, 158962. [Google Scholar] [CrossRef]
- Kleypas, J.A.; Langdon, C. Coral reefs and changing seawater carbonate chemistry. Coral Reefs Clim. Chang. Sci. Manag. 2006, 61, 73–110. [Google Scholar]
- Andersson, A.J.; Yeakel, K.L.; Bates, N.R.; De Putron, S.J. Partial offsets in ocean acidification from changing coral reef biogeochemistry. Nat. Clim. Chang. 2014, 4, 56–61. [Google Scholar] [CrossRef]
- Jokiel, P.L.; Jury, C.P.; Kuffner, I.B. Coral calcification and ocean acidification. Coral Reefs Crossroads 2016, 6, 7–45. [Google Scholar]
- Benson, B.B.; Krause, D. The concentration and isotopic fractionation of oxygen dissolved in fresh water and seawater in equilibrium with the atmosphere. Limnol. Oceanogr. 1984, 29, 620–632. [Google Scholar] [CrossRef]
- Redfield, A.C. The biological control of chemical factors in the environment. Am. Sci. 1958, 46, 230A, 205–221. [Google Scholar]
- Cantoni, C.; Luchetta, A.; Celio, M.; Cozzi, S.; Raicich, F.; Catalano, G. Carbonate system variability in the Gulf of Trieste (North Adriatic Sea). Estuar. Coast. Shelf Sci. 2012, 115, 51–62. [Google Scholar] [CrossRef]
- Leclercq, N.; Gattuso, J.P.; Jaubert, J. Primary production, respiration, and calcification of a coral reef mesocosm under increased CO2 partial pressure. Limnol. Oceanogr. 2002, 47, 558–564. [Google Scholar] [CrossRef]
- Middelburg, J.J.; Duarte, C.M.; Gattuso, J.P. Respiration in coastal benthic communities. In Respiration in Aquatic Ecosystems; Oxford University Press: Oxford, UK, 2005; pp. 206–224. [Google Scholar]
- Ulstrup, K.E.; Kühl, M.; Van Oppen, M.J.; Cooper, T.F.; Ralph, P.J. Variation in photosynthesis and respiration in geographically distinct populations of two reef-building coral species. Aquat. Biol. 2011, 12, 241–248. [Google Scholar] [CrossRef]
- Khripounoff, A.; Caprais, J.C.; Le Bruchec, J.; Rodier, P.; Noel, P.; Cathalot, C. Deep cold-water coral ecosystems in the Brittany submarine canyons (Northeast Atlantic): Hydrodynamics, particle supply, respiration, and carbon cycling. Limnol. Oceanogr. 2014, 59, 87–98. [Google Scholar] [CrossRef]
- Bythell, J.C.; Wild, C. Biology and ecology of coral mucus release. J. Exp. Mar. Biol. Ecol. 2011, 408, 88–93. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Sampling Time | TA μmol kg−1 | DIC μmol kg−1 | pHT | pCO2 μatm | ΩA | Reference |
|---|---|---|---|---|---|---|---|
| Pedra da Risca do Meio Coral Reef | August and November 2020 | 2325 ± 19 | 2019 ± 16 | 7.980 ± 0.008 | 475 ± 28 | n.d. | Cotovicz et al. [30] |
| Great Barrier Reef–Australia | September 2009 to August 2016 | 2288 ± 44 | 1989 ± 45 | 8.03 ± 0.05 | 404 ± 46 | n.d. | Lønborg et al. [55] |
| Trawler Reef | August 2014 | 2289.3 ± 4.9 | 2003.0 ± 19.7 | 8.011 ± 0.020 | n.d. | n.d. | Hannan et al. [56] |
| Big Vicki’s Reef | August 2014 | 2284.0 ± 9.8 | 1984.7 ± 14.7 | 8.044 ± 0.023 | n.d. | n.d. | Hannan et al. [56] |
| Palfrey Reef | August 2014 | 2278.4 ± 7.4 | 1981.9 ± 22.7 | 8.048 ± 0.036 | n.d. | n.d. | Hannan et al. [56] |
| Coral Reef Lagoon Kaneohe Bay | September 2003 to September 2004 | 2180 ± 36 | 1920 ± 16 | n.d. | 460 ± 52 | n.d. | Fagan and Mackenzie [51] |
| The coast of Iriomote Island (Japan) | August 2017 | 2211 ± 44 | 1878 ± 103 | 8.009 ± 0.177 | 415 ± 81 | n.d. | Akhand et al. [52] |
| Reef flat in Northeastern Brazil | July 2006 | 1857.6 ± 42.1 | 1623.0 ± 39.2 | 8.13 ± 0.12 | 371.6 ± 9.2 | n.d. | Akhand et al. [52] |
| Luhuitou fringing reef, Sanya Bay, China | July 2010 | 2312.1 ± 15.3 | 1994.7 ± 40.9 | 8.007 ± 0.048 | 420.1 ± 62.4 | n.d. | Zhang et al. [53] |
| Yongxing Island, China | July to August 2009 | 2421 ± 142 | n.d. | 8.23 ± 0.22 | 456 ± 249 | n.d. | Yan et al. [54] |
| Fiery Cross Reef, China | July to August 2009 | 2240 ± 56 | n.d. | 8.22 ± 0.03 | 395 ± 25 | n.d. | Yan et al. [54] |
| Great Barrier Reef, Australia | November 2011 to April 2012 | 2276 ± 16 | 1954 ± 25 | 8.03 ± 0.03 | 404 ± 40 | n.d. | Albright et al. [57] |
| Gulf of California, Mexico | January to May 2014 | n.d. | n.d. | n.d. | n.d. | 3.03 ± 0.05 | Norzagaray et al. [58] |
| Reef flats of Kiritimati Island | May and June 2018 | 2256.1 ± 22.2 | 1989.8 ± 59.6 | 7.95 ± 0.08 | n.d. | 3.1 ± 0.4 | Knebel et al. [59] |
| Dongshan coral habitat | May 2019 | 2221 ± 0 | 1960 ± 4 | 402 ± 6 | 2.92 ± 0.05 | Dong et al. [31] | |
| August 2019 | 2249 ± 1 | 2038 ± 0 | 619 ± 15 | 2.47 ± 0.01 | |||
| December 2019 | 2209 ± 1 | 2009 ± 4 | 436 ± 11 | 2.26 ± 0.03 | |||
| Gray’s Reef | July 2006 to October 2007 | n.d. | n.d. | 8.08 ± 0.05 | 377 ± 10 | 3.47 ± 0.46 | Xue et al. [42,43] |
| Eastern coast of Shenzhen, China | November 2022 | 2184.6 ± 24.6 | 1879.7 ± 30.5 | 8.12 ± 0.03 | 317.8 ± 26.2 | 3.40 ± 0.18 | In this study |
| February 2023 | 2219.6 ± 13.8 | 1998.4 ± 15.9 | 8.17 ± 0.03 | 405.2 ±35.8 | 2.50 ± 0.11 | ||
| May 2023 | 2178.8 ± 21.7 | 1938.3 ± 22.7 | 8.12 ± 0.03 | 458.5 ± 33.7 | 2.81 ± 0.15 | ||
| August 2023 | 2153.7 ± 37.0 | 1914.8 ± 58.5 | 8.06 ± 0.07 | 579.5 ± 119.8 | 2.89 ± 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Zhang, Z.; Xie, Z.; Chen, B.; Zheng, H.; Liao, B.; Zhou, J.; Xiao, B. Seasonal Controls of Seawater CO2 Systems in Subtropical Coral Reefs: A Case Study from the Eastern Coast of Shenzhen, China. Water 2023, 15, 4124. https://doi.org/10.3390/w15234124
Yang B, Zhang Z, Xie Z, Chen B, Zheng H, Liao B, Zhou J, Xiao B. Seasonal Controls of Seawater CO2 Systems in Subtropical Coral Reefs: A Case Study from the Eastern Coast of Shenzhen, China. Water. 2023; 15(23):4124. https://doi.org/10.3390/w15234124
Chicago/Turabian StyleYang, Bo, Zhuo Zhang, Ziqiang Xie, Bogui Chen, Huina Zheng, Baolin Liao, Jin Zhou, and Baohua Xiao. 2023. "Seasonal Controls of Seawater CO2 Systems in Subtropical Coral Reefs: A Case Study from the Eastern Coast of Shenzhen, China" Water 15, no. 23: 4124. https://doi.org/10.3390/w15234124