

Effects of Short-Term Sodium Nitrate versus Sodium Chloride Supplementation on Energy and Lipid Metabolism during High-Intensity Intermittent Exercise in Athletes
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
2.3. Supplementation
2.4. Measurements
2.5. Data Analysis, Presentation, and Statistics
3. Results
4. Discussion
4.1. Methodological Considerations—Nitrate Dosage, Oxygen Uptake, and Measurements
4.2. Oxygen Uptake
4.3. Glucose Metabolism and Respiratory Exchange Ratio
4.4. Lipid Metabolism
4.5. Metabolites of Energy-Rich Phosphates
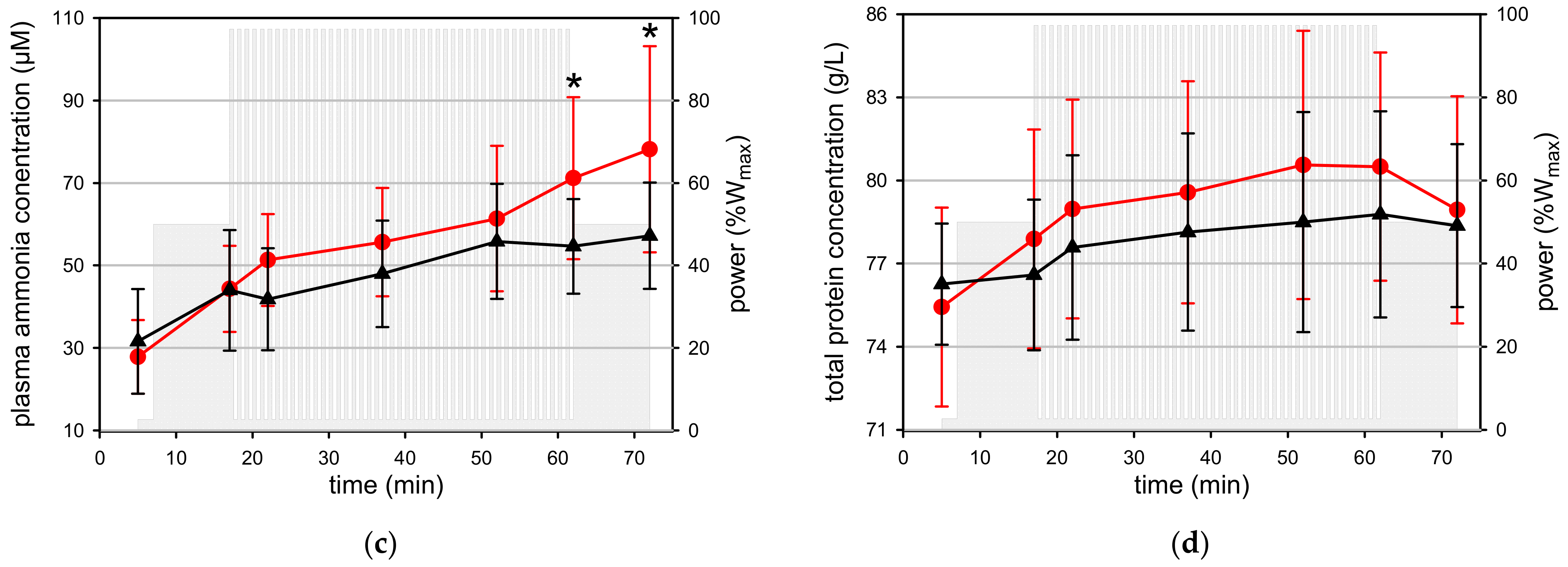
4.6. Amino Acid Metabolism—Ammonia
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Domínguez, R.; Cuenca, E.; Maté-Muñoz, J.; García-Fernández, P.; Serra-Paya, N.; Estevan, M.; Herreros, P.; Garnacho-Castaño, M. Effects of Beetroot Juice Supplementation on Cardiorespiratory Endurance in Athletes. A Systematic Review. Nutrients 2017, 9, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Macuh, M.; Knap, B. Effects of Nitrate Supplementation on Exercise Performance in Humans: A Narrative Review. Nutrients 2021, 13, 3183. [Google Scholar] [CrossRef]
- Van De Walle, G.P.; Vukovich, M.D. The Effect of Nitrate Supplementation on Exercise Tolerance and Performance: A Systematic Review and Meta-Analysis. J. Strength Cond. Res. 2018, 32, 1796–1808. [Google Scholar] [CrossRef] [PubMed]
- Hoon, M.W.; Johnson, N.A.; Chapman, P.G.; Burke, L.M. The Effect of Nitrate Supplementation on Exercise Performance in Healthy Individuals: A Systematic Review and Meta-Analysis. Int. J. Sport. Nutr. Exerc. Metab. 2013, 23, 522–532. [Google Scholar] [CrossRef] [Green Version]
- McMahon, N.F.; Leveritt, M.D.; Pavey, T.G. The Effect of Dietary Nitrate Supplementation on Endurance Exercise Performance in Healthy Adults: A Systematic Review and Meta-Analysis. Sport. Med. 2017, 47, 735–756. [Google Scholar] [CrossRef] [Green Version]
- Pawlak-Chaouch, M.; Boissi Ere, J.; Gamelin, F.X.; Egory Cuvelier, G.; Berthoin, S.; Aucouturier, J.; Boissière, J.; Gamelin, F.X.; Cuvelier, G.; Berthoin, S.; et al. Effect of Dietary Nitrate Supplementation on Metabolic Rate during Rest and Exercise in Human: A Systematic Review and a Meta-Analysis. Nitric. Oxide 2016, 53, 65–76. [Google Scholar] [CrossRef]
- Ignarro, L.J. Biosynthesis and Metabolism of Endothelium-Derived Nitric Oxide. Annu. Rev. Pharmacol. Toxicol. 1990, 30, 535–560. [Google Scholar] [CrossRef] [PubMed]
- Nisoli, E.; Carruba, M.O. Nitric Oxide and Mitochondrial Biogenesis. J. Cell. Sci. 2006, 119, 2855–2862. [Google Scholar] [CrossRef] [Green Version]
- Rubanyi, G.M. Endothelium-derived Relaxing and Contracting Factors. J. Cell. Biochem. 1991, 46, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Bradley, S.J.; Kingwell, B.A.; McConell, G.K. Nitric Oxide Synthase Inhibition Reduces Leg Glucose Uptake but Not Blood Flow during Dynamic Exercise in Humans. Diabetes 1999, 48, 1815–1821. [Google Scholar] [CrossRef]
- Lundberg, J.O.; Carlström, M.; Weitzberg, E. Metabolic Effects of Dietary Nitrate in Health and Disease. Cell. Metab. 2018, 28, 9–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nisoli, E.; Clementi, E.; Paolucci, C.; Cozzi, V.; Tonello, C.; Sciorati, C.; Bracale, R.; Valerio, A.; Francolini, M.; Moncada, S.; et al. Mitochondrial Biogenesis in Mammals: The Role of Endogenous Nitric Oxide. Science 2003, 299, 896–899. [Google Scholar] [CrossRef] [PubMed]
- Larsen, F.J.; Schiffer, T.A.; Borniquel, S.; Sahlin, K.; Ekblom, B.; Lundberg, J.O.; Weitzberg, E. Dietary Inorganic Nitrate Improves Mitochondrial Efficiency in Humans. Cell. Metab. 2011, 13, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Whitfield, J.; Ludzki, A.; Heigenhauser, G.J.F.; Senden, J.M.G.; Verdijk, L.B.; van Loon, L.J.C.; Spriet, L.L.; Holloway, G.P. Beetroot Juice Supplementation Reduces Whole Body Oxygen Consumption but Does Not Improve Indices of Mitochondrial Efficiency in Human Skeletal Muscle. J. Physiol. 2016, 594, 421–435. [Google Scholar] [CrossRef] [Green Version]
- Bailey, S.J.; Fulford, J.; Vanhatalo, A.; Winyard, P.G.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Benjamin, N.; Jones, A.M. Dietary Nitrate Supplementation Enhances Muscle Contractile Efficiency during Knee-Extensor Exercise in Humans. J. Appl. Physiol. 2010, 109, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bailey, S.J.; Winyard, P.; Vanhatalo, A.; Blackwell, J.R.; DiMenna, F.J.; Wilkerson, D.P.; Tarr, J.; Benjamin, N.; Jones, A.M. Dietary Nitrate Supplementation Reduces the O2 Cost of Low-Intensity Exercise and Enhances Tolerance to High-Intensity Exercise in Humans. J. Appl. Physiol. 2009, 107, 1144–1155. [Google Scholar] [CrossRef] [Green Version]
- Lansley, K.E.; Winyard, P.G.; Bailey, S.J.; Vanhatalo, A.; Wilkerson, D.P.; Blackwell, J.R.; Gilchrist, M.; Benjamin, N.; Jones, A.M. Acute Dietary Nitrate Supplementation Improves Cycling Time Trial Performance. Med. Sci. Sport. Exerc. 2011, 43, 1125–1131. [Google Scholar] [CrossRef] [Green Version]
- Larsen, F.J.; Weitzberg, E.; Lundberg, J.O.; Ekblom, B. Effects of Dietary Nitrate on Oxygen Cost during Exercise. Acta Physiol. 2007, 191, 59–66. [Google Scholar] [CrossRef]
- Larsen, F.J.; Weitzberg, E.; Lundberg, J.O.; Ekblom, B. Dietary Nitrate Reduces Maximal Oxygen Consumption While Maintaining Work Performance in Maximal Exercise. Free Radic. Biol. Med. 2010, 48, 342–347. [Google Scholar] [CrossRef]
- Bailey, S.J.; Varnham, R.L.; DiMenna, F.J.; Breese, B.C.; Wylie, L.J.; Jones, A.M. Inorganic Nitrate Supplementation Improves Muscle Oxygenation, O 2 Uptake Kinetics, and Exercise Tolerance at High but Not Low Pedal Rates. J. Appl. Physiol. 2015, 118, 1396–1405. [Google Scholar] [CrossRef]
- Bescús, R.; Rodríguez, F.A.; Iglesias, X.; Ferrer, M.D.; Iborra, E.; Pons, A. Acute Administration of Inorganic Nitrate Reduces V˙O2peak in Endurance Athletes. Med. Sci. Sport. Exerc. 2011, 43, 1979–1986. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, J.; Vanhatalo, A.; Wilkerson, D.P.; Wylie, L.J.; Jones, A.M. Effects of Nitrate on the Power-Duration Relationship for Severe-Intensity Exercise. Med. Sci. Sport. Exerc. 2013, 45, 1798–1806. [Google Scholar] [CrossRef] [Green Version]
- Breese, B.C.; Mcnarry, M.A.; Marwood, S.; Blackwell, J.R.; Bailey, S.J.; Jones, A.M. Beetroot Juice Supplementation Speeds O2 Uptake Kinetics and Improves Exercise Tolerance during Severe-Intensity Exercise Initiated from an Elevated Metabolic Rate. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 305, R1441–R1450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashmore, T.; Roberts, L.D.; Morash, A.J.; Kotwica, A.O.; Finnerty, J.; West, J.A.; Murfitt, S.A.; Fernandez, B.O.; Branco, C.; Cowburn, A.S.; et al. Nitrate Enhances Skeletal Muscle Fatty Acid Oxidation via a Nitric Oxide-CGMP-PPAR-Mediated Mechanism. BMC Biol. 2015, 13, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, A.; Lüersen, K.; Schultheiß, G.; De Pascuallteresa, S.; Mereu, A.; Ipharraguerre, I.R.; Rimbach, G. Supplementation with Nitrate Only Modestly Affects Lipid and Glucose Metabolism in Genetic and Dietaryyinduced Murine Models of Obesity. J. Clin. Biochem. Nutr. 2020, 66, 24–35. [Google Scholar] [CrossRef] [Green Version]
- Tan, R.; Wylie, L.J.; Thompson, C.; Blackwell, J.R.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Beetroot Juice Ingestion during Prolonged Moderate-Intensity Exercise Attenuates Progressive Rise in O2 Uptake. J. Appl. Physiol. 2018, 124, 1254–1263. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.; Vanhatalo, A.; Jell, H.; Fulford, J.; Carter, J.; Nyman, L.; Bailey, S.J.; Jones, A.M. Dietary Nitrate Supplementation Improves Sprint and High-Intensity Intermittent Running Performance. Nitric. Oxide 2016, 61, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.; Vanhatalo, A.; Kadach, S.; Wylie, L.J.; Fulford, J.; Ferguson, S.K.; Blackwell, J.R.; Bailey, S.J.; Jones, A.M. Discrete Physiological Effects of Beetroot Juice and Potassium Nitrate Supplementation Following 4-Wk Sprint Interval Training. J. Appl. Physiol. 2018, 124, 1519–1528. [Google Scholar] [CrossRef]
- Muggeridge, D.J.; Sculthorpe, N.; James, P.E.; Easton, C. The Effects of Dietary Nitrate Supplementation on the Adaptations to Sprint Interval Training in Previously Untrained Males. J. Sci. Med. Sport. 2017, 20, 92–97. [Google Scholar] [CrossRef] [Green Version]
- Wong, T.H.; Sim, A.; Burns, S.F. The Effect of Beetroot Ingestion on High-Intensity Interval Training: A Systematic Review and Meta-Analysis. Nutrients 2021, 13, 3674. [Google Scholar] [CrossRef]
- Anderson, O.K.; Martinez-Ferran, M.; Lorenzo-Calvo, J.; Jiménez, S.L.; Pareja-Galeano, H. Effects of Nitrate Supplementation on Muscle Strength and Mass: A Systematic Review. J. Strength Cond. Res. 2022, 36, 3562–3570. [Google Scholar] [CrossRef] [PubMed]
- Delleli, S.; Ouergui, I.; Messaoudi, H.; Trabelsi, K.; Glenn, J.M.; Ammar, A.; Chtourou, H. Does Beetroot Supplementation Improve Performance in Combat Sports Athletes? A Systematic Review of Randomized Controlled Trials. Nutrients 2023, 15, 398. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, A.M.; Townsend, J.R.; Pinzone, A.G.; Hoffman, J.R.; Eckerson, J.M.; Gonzalez, A.M.; Townsend, J.R.; Pinzone, A.G.; Hoffman, J.R. Supplementation with Nitric Oxide Precursors for Strength Performance: A Review of the Current Literature. Nutrients 2023, 15, 660. [Google Scholar] [CrossRef] [PubMed]
- Wong, T.H.; Sim, A.; Burns, S.F. The Effects of Nitrate Ingestion on High-Intensity Endurance Time-Trial Performance: A Systematic Review and Meta-Analysis. J. Exerc. Sci. Fit. 2022, 20, 305–316. [Google Scholar] [CrossRef]
- Finkel, A.; Röhrich, M.A.; Maassen, N.; Lützow, M.; Blau, L.S.; Hanff, E.; Tsikas, D.; Maassen, M. Long-Term Effects of NO3− on the Relationship between Oxygen Uptake and Power after Three Weeks of Supplemented HIHVT. J. Appl. Physiol. 2018, 125, 1997–2007. [Google Scholar] [CrossRef]
- Hanff, E. Einfluss von Leistungssport Auf Die Blutkonzentration von Homoarginin, Guanidinoacetat Und Arginin: Analytik, Biochemie, Physiologie Und Pharmakologie. Bachelor’s Thesis, Medizinische Hochschule Hannover, Hannover, Germany, 2015. [Google Scholar]
- Domínguez, R.; Maté-Muñoz, J.L.; Cuenca, E.; García-Fernández, P.; Mata-Ordoñez, F.; Lozano-Estevan, M.C.; Veiga-Herreros, P.; da Silva, S.F.; Garnacho-Castaño, M.V. Effects of Beetroot Juice Supplementation on Intermittent High-Intensity Exercise Efforts. J. Int. Soc. Sport. Nutr. 2018, 15, 2. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D.; Maassen, N.; Thorns, A.; Finkel, A.; Lützow, M.; Röhrig, M.A.; Blau, L.S.; Dimina, L.; Mariotti, F.; Beckmann, B.; et al. Short-Term Supplementation of Sodium Nitrate vs. Sodium Chloride Increases Homoarginine Synthesis in Young Men Independent of Exercise. Int. J. Mol. Sci. 2022, 23, 649. [Google Scholar] [CrossRef]
- Schrader, M.; Treff, B.; Sandholtet, T.; Maassen, N.; Shushakov, V.; Kaesebieter, J.; Maassen, M. Carbohydrate Supplementation Stabilises Plasma Sodium during Training with High Intensity. Eur. J. Appl. Physiol. 2016, 116, 1841–1853. [Google Scholar] [CrossRef] [PubMed]
- Maassen, N.; Busse, M.W. The Relationship between Lactic Acid and Work Load: A Measure for Endurance Capacity or an Indicator of Carbohydrate Deficiency? Eur. J. Appl. Physiol. Occup. Physiol. 1989, 58, 728–737. [Google Scholar] [CrossRef]
- Frische, M. Die Wirkung Eines Hochintensiven, Intervallartigen Trainings Auf Die Leistungs- Und Regenerationsfähigkeit. Ph.D. Thesis, Gottfried Wilhelm Leibniz Universität Hannover, Hannover, Germany, 2007. [Google Scholar]
- Hanff, E.; Eisenga, M.F.; Beckmann, B.; Bakker, S.J.L.; Tsikas, D. Simultaneous Pentafluorobenzyl Derivatization and GC-ECNICI-MS Measurement of Nitrite and Malondialdehyde in Human Urine: Close Positive Correlation between These Disparate Oxidative Stress Biomarkers. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2017, 1043, 167–175. [Google Scholar] [CrossRef]
- Tsikas, D. Simultaneous Derivatization and Quantification of the Nitric Oxide Metabolites Nitrite and Nitrate in Biological Fluids by Gas Chromatography/Mass Spectrometry. Anal. Chem. 2000, 72, 4064–4072. [Google Scholar] [CrossRef] [PubMed]
- Maughan, R.J.; Burke, L.M.; Dvorak, J.; Larson-Meyer, D.E.; Peeling, P.; Phillips, S.M.; Rawson, E.S.; Walsh, N.P.; Garthe, I.; Geyer, H.; et al. IOC Consensus Statement: Dietary Supplements and the High-Performance Athlete. Br. J. Sport. Med. 2018, 52, 439–455. [Google Scholar] [CrossRef] [PubMed]
- Shannon, O.M.; Allen, J.D.; Bescos, R.; Burke, L.; Clifford, T.; Easton, C.; Gonzalez, J.T.; Jones, A.M.; Jonvik, K.L.; Larsen, F.J.; et al. Dietary Inorganic Nitrate as an Ergogenic Aid: An Expert Consensus Derived via the Modified Delphi Technique. Sport. Med. 2022, 52, 2537–2558. [Google Scholar] [CrossRef] [PubMed]
- Flueck, J.L.; Bogdanova, A.; Mettler, S.; Perret, C. Is Beetroot Juice More Effective than Sodium Nitrate? The Effects of Equimolar Nitrate Dosages of Nitrate-Rich Beetroot Juice and Sodium Nitrate on Oxygen Consumption during Exercise. Appl. Physiol. Nutr. Metab. 2016, 41, 421–429. [Google Scholar] [CrossRef]
- Flueck, J.L.; Perret, C.; Gallo, A.; Moelijker, N.; Bogdanov, N.; Bogdanova, A. Influence of Equimolar Doses of Beetroot Juice and Sodium Nitrate on Time Trial Performance in Handcycling. Nutrients 2019, 11, 1642. [Google Scholar] [CrossRef] [Green Version]
- Wylie, L.J.; Park, J.W.; Vanhatalo, A.; Kadach, S.; Black, M.I.; Stoyanov, Z.; Schechter, A.N.; Jones, A.M.; Piknova, B. Human Skeletal Muscle Nitrate Store: Influence of Dietary Nitrate Supplementation and Exercise. J. Physiol. 2019, 597, 5565–5576. [Google Scholar] [CrossRef] [Green Version]
- Piknova, B.; Schechter, A.N.; Park, J.W.; Vanhatalo, A.; Jones, A.M. Skeletal Muscle Nitrate as a Regulator of Systemic Nitric Oxide Homeostasis. Exerc. Sport. Sci. Rev. 2022, 50, 2–13. [Google Scholar] [CrossRef]
- Piknova, B.; Park, J.W.; Kwan Lam, K.; Schechter, A.N. Nitrate as a Source of Nitrite and Nitric Oxide during Exercise Hyperemia in Rat Skeletal Muscle. Nitric Oxide 2016, 55–56, 54–61. [Google Scholar] [CrossRef] [Green Version]
- Kadach, S.; Piknova, B.; Black, M.I.; Park, J.W.; Wylie, L.J.; Stoyanov, Z.; Thomas, S.M.; McMahon, N.F.; Vanhatalo, A.; Schechter, A.N.; et al. Time Course of Human Skeletal Muscle Nitrate and Nitrite Concentration Changes Following Dietary Nitrate Ingestion. Nitric Oxide 2022, 121, 1–10. [Google Scholar] [CrossRef]
- Porcelli, S.; Ramaglia, M.; Bellistri, G.; Pavei, G.; Pugliese, L.; Montorsi, M.; Rasica, L.; Marzorati, M. Aerobic Fitness Affects the Exercise Performance Responses to Nitrate Supplementation. Med. Sci. Sport. Exerc. 2015, 47, 1643–1651. [Google Scholar] [CrossRef] [Green Version]
- Boorsma, R.K.; Whitfield, J.; Spriet, L.L. Beetroot Juice Supplementation Does Not Improve Performance of Elite 1500-m Runners. Med. Sci. Sport. Exerc. 2014, 46, 2326–2334. [Google Scholar] [CrossRef] [PubMed]
- Christensen, P.M.; Nyberg, M.; Bangsbo, J. Influence of Nitrate Supplementation on VO2 Kinetics and Endurance of Elite Cyclists. Scand. J. Med. Sci. Sport. 2013, 23, e21–e31. [Google Scholar] [CrossRef] [PubMed]
- Wilkerson, D.P.; Hayward, G.M.; Bailey, S.J.; Vanhatalo, A.; Blackwell, J.R.; Jones, A.M. Influence of Acute Dietary Nitrate Supplementation on 50 Mile Time Trial Performance in Well-Trained Cyclists. Eur. J. Appl. Physiol. 2012, 112, 4127–4134. [Google Scholar] [CrossRef] [PubMed]
- Wylie, L.J.; Mohr, M.; Krustrup, P.; Jackman, S.R.; Ermιdis, G.; Kelly, J.; Black, M.I.; Bailey, S.J.; Vanhatalo, A.; Jones, A.M. Dietary Nitrate Supplementation Improves Team Sport-Specific Intense Intermittent Exercise Performance. Eur. J. Appl. Physiol. 2013, 113, 1673–1684. [Google Scholar] [CrossRef]
- Dreißigacker, U.; Wendt, M.; Wittke, T.; Tsikas, D.; Maassen, N. Positive Correlation between Plasma Nitrite and Performance during High-Intensive Exercise but Not Oxidative Stress in Healthy Men. Nitric Oxide 2010, 23, 128–135. [Google Scholar] [CrossRef]
- Haghikia, A.; Yanchev, G.R.; Kayacelebi, A.A.; Hanff, E.; Bledau, N.; Widera, C.; Sonnenschein, K.; Haghikia, A.; Weissenborn, K.; Bauersachs, J.; et al. The Role of L-Arginine/l-Homoarginine/Nitric Oxide Pathway for Aortic Distensibility and Intima-Media Thickness in Stroke Patients. Amino Acids 2017, 49, 1111–1121. [Google Scholar] [CrossRef]
- Hanff, E.; Lützow, M.; Kayacelebi, A.A.; Finkel, A.; Maassen, M.; Yanchev, G.R.; Haghikia, A.; Bavendiek, U.; Buck, A.; Lücke, T.; et al. Simultaneous GC-ECNICI-MS Measurement of Nitrite, Nitrate and Creatinine in Human Urine and Plasma in Clinical Settings. J. Chromatogr. B 2017, 1047, 207–214. [Google Scholar] [CrossRef]
- Jakubcik, E.M.; Rutherfurd-Markwick, K.; Chabert, M.; Wong, M.; Ali, A. Pharmacokinetics of Nitrate and Nitrite Following Beetroot Juice Drink Consumption. Nutrients 2021, 13, 281. [Google Scholar] [CrossRef]
- Essén, B.; Kaijser, L. Regulation of Glycolysis in Intermittent Exercise in Man. J. Physiol. 1978, 281, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Essén, B.; Hagenfeldt, L.; Kaijser, L. Utilization of Blood-Borne and Intramuscular Substrates during Continuous and Intermittent Exerise in Men. J. Sport. Med. 1977, 265, 489–506. [Google Scholar] [CrossRef]
- Eigendorf, J.; Maassen, M.; Apitius, D.; Maassen, N. Energy Metabolism in Continuous, High-Intensity, and Sprint Interval Training Protocols with Matched Mean Intensity. J. Strength Cond. Res. 2021, 35, 3104–3110. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.; Wylie, L.J.; Blackwell, J.R.; Fulford, J.; Black, M.I.; Kelly, J.; McDonagh, S.T.J.; Carter, J.; Bailey, S.J.; Vanhatalo, A.; et al. Influence of Dietary Nitrate Supplementation on Physiological and Muscle Metabolic Adaptations to Sprint Interval Training. J. Appl. Physiol. 2017, 122, 642–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, L.J.; Bailey, S.J.; Kelly, J.; Blackwell, J.R.; Vanhatalo, A.; Jones, A.M. Influence of Beetroot Juice Supplementation on Intermittent Exercise Performance. Eur. J. Appl. Physiol. 2016, 116, 415–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyakayiru, J.; Jonvik, K.L.; Trommelen, J.; Pinckaers, P.J.M.; Senden, J.M.; van Loon, L.J.C.; Verdijk, L.B. Beetroot Juice Supplementation Improves High-Intensity Intermittent Type Exercise Performance in Trained Soccer Players. Nutrients 2017, 9, 314. [Google Scholar] [CrossRef] [Green Version]
- Esen, O.; Domínguez, R.; Karayigit, R. Acute Beetroot Juice Supplementation Enhances Intermittent Running Performance but Does Not Reduce Oxygen Cost of Exercise among Recreational Adults. Nutrients 2022, 14, 2839. [Google Scholar] [CrossRef]
- Khalifi, S.; Rahimipour, A.; Jeddi, S.; Ghanbari, M.; Kazerouni, F.; Ghasemi, A. Dietary Nitrate Improves Glucose Tolerance and Lipid Profile in an Animal Model of Hyperglycemia. Nitric Oxide 2015, 44, 24–30. [Google Scholar] [CrossRef]
- Li, T.; Lu, X.; Sun, Y.; Yang, X. Effects of Spinach Nitrate on Insulin Resistance, Endothelial Dysfunction Markers and Inflammation in Mice with High-Fat and High-Fructose Consumption. Food Nutr. Res. 2016, 60, 32010. [Google Scholar] [CrossRef] [Green Version]
- Lai, Y.-C.; Tabima, D.M.; Dube, J.J.; Hughan, K.S.; Vanderpool, R.R.; Goncharov, D.A.; St. Croix, C.M.; Garcia-Ocaña, A.; Goncharova, E.A.; Tofovic, S.P.; et al. SIRT3–AMP-Activated Protein Kinase Activation by Nitrite and Metformin Improves Hyperglycemia and Normalizes Pulmonary Hypertension Associated with Heart Failure with Preserved Ejection Fraction. Circulation 2016, 133, 717–731. [Google Scholar] [CrossRef]
- Jiang, H.; Torregrossa, A.C.; Potts, A.; Pierini, D.; Aranke, M.; Garg, H.K.; Bryan, N.S. Dietary Nitrite Improves Insulin Signaling through GLUT4 Translocation. Free Radic. Biol. Med. 2014, 67, 51–57. [Google Scholar] [CrossRef]
- Gheibi, S.; Bakhtiarzadeh, F.; Jeddi, S.; Farrokhfall, K.; Zardooz, H.; Ghasemi, A. Nitrite Increases Glucose-Stimulated Insulin Secretion and Islet Insulin Content in Obese Type 2 Diabetic Male Rats. Nitric Oxide 2017, 64, 39–51. [Google Scholar] [CrossRef]
- Larsen, F.J.; Schiffer, T.A.; Ekblom, B.; Mattsson, M.P.; Checa, A.; Wheelock, C.E.; Nyström, T.; Lundberg, J.O.; Weitzberg, E. Dietary Nitrate Reduces Resting Metabolic Rate: A Randomized, Crossover Study in Humans. Am. J. Clin. Nutr. 2014, 99, 843–850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wylie, L.J.; Kelly, J.; Bailey, S.J.; Blackwell, J.R.; Skiba, P.F.; Winyard, P.G.; Jeukendrup, A.E.; Vanhatalo, A.; Jones, A.M. Beetroot Juice and Exercise: Pharmacodynamic and Dose-Response Relationships. J. Appl. Physiol. 2013, 115, 325–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McNally, B.D.; Moran, A.; Watt, N.T.; Ashmore, T.; Whitehead, A.; Murfitt, S.A.; Kearney, M.T.; Cubbon, R.M.; Murray, A.J.; Griffin, J.L.; et al. Inorganic Nitrate Promotes Glucose Uptake and Oxidative Catabolism in White Adipose Tissue Through the XOR-Catalyzed Nitric Oxide Pathway. Diabetes 2020, 69, 893–901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haskell, W.L. The Influence of Exercise on the Concentration of Triglyceride and Cholesterol in Human Plasma. Exerc. Sport. Sci. Rev. 1984, 12, 205–244. [Google Scholar] [CrossRef] [PubMed]
- Hurley, B.F.; Nemeth, P.M.; Martin, W.H.; Hagberg, J.M.; Dalsky, G.P.; Holloszy, J.O. Muscle Triglyceride Utilization during Exercise: Effect of Training. J. Appl. Physiol. 1986, 60, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Ranallo, R.F.; Rhodes, E.C. Lipid Metabolism During Exercise. Sport. Med. 1998, 26, 29–42. [Google Scholar] [CrossRef]
- Talanian, J.L.; Galloway, S.D.R.R.; Heigenhauser, G.J.F.F.; Bonen, A.; Spriet, L.L. Two Weeks of High-Intensity Aerobic Interval Training Increases the Capacity for Fat Oxidation during Exercise in Women. J. Appl. Physiol. 2007, 102, 1439–1447. [Google Scholar] [CrossRef]
- Roberts, L.D.; Ashmore, T.; Kotwica, A.O.; Murfitt, S.A.; Fernandez, B.O.; Feelisch, M.; Murray, A.J.; Griffin, J.L. Inorganic Nitrate Promotes the Browning of White Adipose Tissue Through the Nitrate-Nitrite-Nitric Oxide Pathway. Diabetes 2015, 64, 471–484. [Google Scholar] [CrossRef] [Green Version]
- Maassen, M. Die Wirkung von Kohlenhydratgaben Während Des Trainings Auf Die Entwicklung Der Ausdauer-, Sprint-Und Regenerationsfähigkeit. Ph.D. Thesis, Leibniz Universität Hannover, Hannover, Germany, 2011. [Google Scholar]
- MacDonald, I. Diet and Triglyceride Metabolism. J. Clin. Pathol. Suppl. Assoc. Clin. Pathol. 1973, 5, 22–25. [Google Scholar] [CrossRef] [Green Version]
- Parks, E.J. Effect of Dietary Carbohydrate on Triglyceride Metabolism in Humans. J. Nutr. 2001, 131, 2772S–2774S. [Google Scholar] [CrossRef] [Green Version]
- Elia, M.; Khan, M.; Calder, G.; Kurpad, A. Glycerol Exchange across the Human Forearm Assessed by a Combination of Tracer and Arteriovenous Exchange Techniques. Clin. Sci. 1993, 84, 99–104. [Google Scholar] [CrossRef]
- Romijn, J.A.; Coyle, E.F.; Sidossis, L.S.; Gastaldelli, A.; Horowitz, J.F.; Endert, E.; Wolfe, R.R. Regulation of Endogenous Fat and Carbohydrate Metabolism in Relation to Exercise Intensity and Duration. Am. J. Physiol. Endocrinol. Metab. 1993, 265, E380–E391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiens, B.; Richter, E.A. Utilization of Skeletal Muscle Triacylglycerol during Postexercise Recovery in Humans. Am. J. Physiol. 1998, 275, E332–E337. [Google Scholar] [CrossRef] [PubMed]
- Hardman, A.E. The Influence of Exercise on Postprandial Triacylglycerol Metabolism. Atherosclerosis 1998, 141, S93–S100. [Google Scholar] [CrossRef] [PubMed]
- Oscai, L.B.; Essig, D.A.; Palmer, W.K. Lipase Regulation of Muscle Triglyceride Hydrolysis. J. Appl. Physiol. 1990, 69, 1571–1577. [Google Scholar] [CrossRef]
- Liu, M.S.; Jirik, F.R.; LeBoeuf, R.C.; Henderson, H.; Castellani, L.W.; Lusis, A.J.; Ma, Y.; Forsythe, I.J.; Zhang, H.; Kirk, E.; et al. Alteration of Lipid Profiles in Plasma of Transgenic Mice Expressing Human Lipoprotein Lipase. J. Biol. Chem. 1994, 269, 11417–11424. [Google Scholar] [CrossRef]
- Shimada, M.; Shimano, H.; Gotoda, T.; Yamamoto, K.; Kawamura, M.; Inaba, T.; Yazaki, Y.; Yamada, N. Overexpression of Human Lipoprotein Lipase in Transgenic Mice. Resistance to Diet-Induced Hypertriglyceridemia and Hypercholesterolemia. J. Biol. Chem. 1993, 268, 17924–17929. [Google Scholar] [CrossRef]
- Budoff, M. Triglycerides and Triglyceride-Rich Lipoproteins in the Causal Pathway of Cardiovascular Disease. Am. J. Cardiol. 2016, 118, 138–145. [Google Scholar] [CrossRef] [Green Version]
- Abdel-Maksoud, M.F.; Hokanson, J.E. The Complex Role of Triglycerides in Cardiovascular Disease. Semin. Vasc. Med. 2002, 2, 325–333. [Google Scholar] [CrossRef]
- Sahlin, K.; Tonkonogi, M.; Söderlund, K. Plasma Hypoxanthine and Ammonia in Humans during Prolonged Exercise. Eur. J. Appl. Physiol. Occup. Physiol. 1999, 80, 417–422. [Google Scholar] [CrossRef]
- Patterson, V.H.; Kaiser, K.K.; Brooke, M.H. Forearm Exercise Increases Plasma Hypoxanthine. J. Neurol. Neurosurg. Psychiatry 1982, 45, 552–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ketai, L.H.; Simon, R.H.; Kreit, J.W.; Grum, C.M. Plasma Hypoxanthine and Exercise. Am. Rev. Respir. Dis. 1987, 136, 98–101. [Google Scholar] [CrossRef] [PubMed]
- Miller, S.G.; Hafen, P.S.; Brault, J.J. Increased Adenine Nucleotide Degradation in Skeletal Muscle Atrophy. Int. J. Mol. Sci. 2020, 21, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battelli, M.G.; Polito, L.; Bortolotti, M.; Bolognesi, A. Xanthine Oxidoreductase-Derived Reactive Species: Physiological and Pathological Effects. Oxid. Med. Cell. Longev. 2016, 2016, 3527579. [Google Scholar] [CrossRef] [Green Version]
- Heunks, L.M.A.; Viña, J.; van Herwaarden, C.L.A.; Folgering, H.T.M.; Gimeno, A.; Dekhuijzen, P.N.R. Xanthine Oxidase Is Involved in Exercise-Induced Oxidative Stress in Chronic Obstructive Pulmonary Disease. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 1999, 277, R1697–R1704. [Google Scholar] [CrossRef]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Bloch, K.; Schoenheimer, R. Studies in Protein Metabolism. J. Biol. Chem. 1939, 131, 111–119. [Google Scholar] [CrossRef]
- Hammouda, O.; Chtourou, H.; Chahed, H.; Ferchichi, S.; Chaouachi, A.; Kallel, C.; Miled, A.; Chamari, K.; Souissi, N. High Intensity Exercise Affects Diurnal Variation of Some Biological Markers in Trained Subjects. Int. J. Sport. Med. 2012, 33, 886–891. [Google Scholar] [CrossRef]
- Linossier, M.T.; Denis, C.; Dormois, D.; Geyssant, A.; Lacour, J.R. Ergometric and Metabolic Adaptation to a 5-s Sprint Training Programme. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 67, 408–414. [Google Scholar] [CrossRef]
- Thorstensson, A.; Sjödin, B.; Karlsson, J. Enzyme Activities and Muscle Strength after “Sprint Training” in Man. Acta Physiol. Scand. 1975, 94, 313–318. [Google Scholar] [CrossRef]
- Hellsten, Y.; Richter, E.A.; Kiens, B.; Bangsbo, J. AMP Deamination and Purine Exchange in Human Skeletal Muscle during and after Intense Exercise. J. Physiol. 1999, 520, 909–920. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.E.; Turcotte, L.P.; Kiens, B.; Richter, E.A. Training and Muscle Ammonia and Amino Acid Metabolism in Humans during Prolonged Exercise. J. Appl. Physiol. 1995, 78, 725–735. [Google Scholar] [CrossRef] [PubMed]
- Hibbs, J.B.; Westenfelder, C.; Taintor, R.; Vavrin, Z.; Kablitz, C.; Baranowski, R.L.; Ward, J.H.; Menlove, R.L.; McMurry, M.P.; Kushner, J.P.; et al. Evidence for Cytokine-Inducible Nitric Oxide Synthesis from L-Arginine in Patients Receiving Interleukin-2 Therapy. J. Clin. Investig. 1992, 89, 867–877. [Google Scholar] [CrossRef] [PubMed]
- Tiso, M.; Schechter, A.N. Nitrate Reduction to Nitrite, Nitric Oxide and Ammonia by Gut Bacteria under Physiological Conditions. PLoS ONE 2015, 10, e0119712. [Google Scholar] [CrossRef] [Green Version]
- Snow, R.J.; Carey, M.F.; Stathis, C.G.; Febbraio, M.A.; Hargreaves, M. Effect of Carbohydrate Ingestion on Ammonia Metabolism during Exercise in Humans. J. Appl. Physiol. 2000, 88, 1576–1580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langfort, J.; Czarnowski, D.; Żendzian-Piotrowska, M.; Zarzeczny, R.; Górski, J. Short-Term Low-Carbohydrate Diet Dissociates Lactate and Ammonia Thresholds in Men. J. Strength Cond. Res. 2004, 18, 260. [Google Scholar]
- Tsikas, D. Homoarginine in Health and Disease. Curr. Opin. Clin. Nutr. Metab. Care 2023, 26, 42–49. [Google Scholar] [CrossRef]
- May, M.; Kayacelebi, A.A.; Batkai, S.; Jordan, J.; Tsikas, D.; Engeli, S. Plasma and Tissue Homoarginine Concentrations in Healthy and Obese Humans. Amino Acids 2015, 47, 1847–1852. [Google Scholar] [CrossRef]
- Mokhaneli, M.C.; Botha-Le Roux, S.; Fourie, C.M.T.; Böger, R.; Schwedhelm, E.; Mels, C.M.C. L-Homoarginine Is Associated with Decreased Cardiovascular- and All-Cause Mortality. Eur. J. Clin. Investig. 2021, 51, e13472. [Google Scholar] [CrossRef]
- März, W.; Meinitzer, A.; Drechsler, C.; Pilz, S.; Krane, V.; Kleber, M.E.; Fischer, J.; Winkelmann, B.R.; Böhm, B.O.; Ritz, E.; et al. Homoarginine, Cardiovascular Risk, and Mortality. Circulation 2010, 122, 967–975. [Google Scholar] [CrossRef]
- Kayacelebi, A.A.; Minović, I.; Hanff, E.; Frenay, A.-R.S.; de Borst, M.H.; Feelisch, M.; van Goor, H.; Bakker, S.J.L.; Tsikas, D. Low Plasma Homoarginine Concentration Is Associated with High Rates of All-Cause Mortality in Renal Transplant Recipients. Amino Acids 2017, 49, 1193–1202. [Google Scholar] [CrossRef] [PubMed]
- Bondonno, C.P.; Dalgaard, F.; Blekkenhorst, L.C.; Murray, K.; Lewis, J.R.; Croft, K.D.; Kyrø, C.; Torp-Pedersen, C.; Gislason, G.; Tjønneland, A.; et al. Vegetable Nitrate Intake, Blood Pressure and Incident Cardiovascular Disease: Danish Diet, Cancer, and Health Study. Eur. J. Epidemiol. 2021, 36, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Shannon, O.M.; Easton, C.; Shepherd, A.I.; Siervo, M.; Bailey, S.J.; Clifford, T. Dietary Nitrate and Population Health: A Narrative Review of the Translational Potential of Existing Laboratory Studies. BMC Sport. Sci. Med. Rehabil. 2021, 13, 65. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.H.; Bondonno, C.P.; Russell, J.; Flood, V.M.; Lewis, J.R.; Croft, K.D.; Woodman, R.J.; Lim, W.H.; Kifley, A.; Wong, G.; et al. Relationship of Dietary Nitrate Intake from Vegetables with Cardiovascular Disease Mortality: A Prospective Study in a Cohort of Older Australians. Eur. J. Nutr. 2019, 58, 2741–2753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sim, M.; Blekkenhorst, L.C.; Bondonno, N.P.; Radavelli-Bagatini, S.; Peeling, P.; Bondonno, C.P.; Magliano, D.J.; Shaw, J.E.; Woodman, R.; Murray, K.; et al. Dietary Nitrate Intake Is Positively Associated with Muscle Function in Men and Women Independent of Physical Activity Levels. J. Nutr. 2021, 151, 1222–1230. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Nitrate Group | Placebo Group |
|---|---|---|
| Number | 9 | 8 |
| Age (y) | 26.1 ± 4.4 | 26.5 ± 3.4 |
| Height (cm) | 184.7 ± 8.1 | 180.5 ± 6.2 |
| Weight (kg) | 78.3 ± 8.0 | 77.2 ± 10.9 |
| relVO2peak (mL/min/kg) | 57.3 ± 5.4 | 56.1 ± 8.8 |
| relWmax (Wmax/kg) | 4.85 ± 0.62 | 4.83 ± 0.91 |
| Time Point | Group | Nitrate (µM) | Nitrite (µM) |
|---|---|---|---|
| Rest | N | 234.3 ± 66.7 * | 1.51 ± 0.39 * |
| P | 61.7 ± 50.4 | 1.16 ± 0.22 | |
| Warm-up | N | 244.4 ± 78.5 * | 1.56 ± 0.29 * |
| P | 70.1 ± 42.0 | 1.25 ± 0.44 | |
| 5th interval | N | 246.4 ± 74.6 * | 1.57 ± 0.28 * |
| P | 64.6 ± 46.7 | 1.21 ± 0.27 | |
| 20th interval | N | 251.3 ± 71.2 * | 1.60 ± 0.24 * |
| P | 68.3 ± 53.1 | 1.13 ± 0.26 | |
| 35th interval | N | 254.3 ± 71.7 * | 1.50 ± 0.21 * |
| P | 65.8 ± 46.1 | 1.12 ± 0.24 | |
| 45th interval | N | 249.2 ± 68.6 * | 1.40 ± 0.23 |
| P | 71.4 ± 44.9 | 1.21 ± 0.28 | |
| Cool-down | N | 256.1 ± 89.9 * | 1.54 ± 0.36 * |
| P | 67.6 ± 47.4 | 1.15 ± 0.20 |
| Time Point | Group | VE (L/min) | petCO2 (mmHg) | pHv (1) | Lacv (mM) | Gluv (mM) |
|---|---|---|---|---|---|---|
| Rest | N | 14.7 ± 5.0 | 35.9 ± 1.9 | 7.36 ± 0.02 | 1.04 ± 0.25 | 4.55 ± 0.71 |
| P | 13.1 ± 3.1 | 32.6 ± 3.1 | 7.34 ± 0.03 | 1.25 ± 0.48 | 4.36 ± 0.62 | |
| Warm-up | N | 66.1 ± 10.3 | 43.2 ± 4.5 | 7.33 ± 0.03 | 2.31 ± 0.49 | 3.93 ± 0.59 |
| P | 68.8 ± 14.8 | 39.8 ± 2.9 | 7.32 ± 0.03 | 2.43 ± 0.58 | 3.83 ± 0.66 | |
| 5th interval | N | 74.2 ± 12.1 | 41.2 ± 5.8 | 7.33 ± 0.03 | 4.03 ± 0.60 | 3.94 ± 0.61 |
| P | 76.8 ± 17.9 | 38.0 ± 3.7 | 7.33 ± 0.03 | 3.89 ± 0.89 | 3.89 ± 0.78 | |
| 20th interval | N | 79.4 ± 13.6 | 37.8 ± 6.2 | 7.35 ± 0.05 | 5.08 ± 0.90 | 4.23 ± 0.64 |
| P | 83.3 ± 21.6 | 35.2 ± 4.0 | 7.34 ± 0.03 | 5.10 ± 1.29 | 4.67 ± 0.44 | |
| 35th interval | N | 84.8 ± 17.8 | 36.7 ± 6.3 | 7.35 ± 0.04 | 5.46 ± 0.95 | 4.40 ± 0.64 |
| P | 88.7 ± 21.9 | 32.8 ± 3.7 | 7.35 ± 0.04 | 5.57 ± 1.34 | 4.90 ± 0.45 | |
| 45th interval | N | 85.0 ± 14.4 | 35.3 ± 7.1 | 7.36 ± 0.04 | 5.76 ± 0.98 | 4.40 ± 0.57 |
| P | 91.8 ± 23.4 | 32.4 ± 4.7 | 7.36 ± 0.04 | 5.85 ± 1.49 | 4.93 ± 0.63 | |
| Cool-down | N | 78.9 ± 17.8 | 37.1 ± 5.7 | 7.39 ± 0.05 | 3.86 ± 0.95 | 4.23 ± 0.60 |
| P | 86.3 ± 20.0 | 34.4 ± 3.9 | 7.36 ± 0.08 | 4.34 ± 1.55 | 4.73 ± 0.81 |
| Time Point | Group | Nav (mM) | Kv (mM) | HCT (%) | Hb (g/dL) |
|---|---|---|---|---|---|
| Rest | N | 140.5 ± 1.6 | 3.96 ± 0.24 | 44.2 ± 2.2 | 15.9 ± 1.2 |
| P | 140.3 ± 1.2 | 4.14 ± 0.21 | 44.0 ± 3.4 | 15.2 ± 0.9 | |
| Warm-up | N | 142.2 ± 1.5 | 4.45 ± 0.28 | 45.1 ± 2.5 | 16.4 ± 1.2 |
| P | 141.8 ± 1.2 | 4.54 ± 0.25 | 44.7 ± 3.1 | 15.5 ± 1.0 | |
| 5th interval | N | 142.7 ± 1.5 | 4.41 ± 0.16 | 45.4 ± 2.5 | 16.5 ± 1.1 |
| P | 142.0 ± 1.1 | 4.56 ± 0.23 | 45.0 ± 3.1 | 15.6 ± 1.0 | |
| 20th interval | N | 142.9 ± 1.3 | 4.49 ± 0.21 | 45.7 ± 2.5 | 16.6 ± 1.0 |
| P | 142.2 ± 0.8 | 4.62 ± 0.23 | 45.6 ± 3.2 | 15.6 ± 1.0 | |
| 35th interval | N | 143.3 ± 1.3 | 4.56 ± 0.16 | 45.8 ± 2.4 | 16.7 ± 1.1 |
| P | 142.4 ± 0.8 | 4.71 ± 0.28 | 45.4 ± 3.5 | 15.7 ± 1.0 | |
| 45th interval | N | 143.4 ± 1.6 | 4.63 ± 0.25 | 45.8 ± 2.4 | 16.7 ± 1.1 |
| P | 142.8 ± 0.7 | 4.80 ± 0.32 | 45.2 ± 3.6 | 15.7 ± 1.1 | |
| Cool-down | N | 142.9 ± 1.9 | 4.88 ± 0.22 | 45.1 ± 2.4 | 16.6 ± 1.1 |
| P | 142.4 ± 0.8 | 5.06 ± 0.36 | 44.8 ± 3.7 | 15.7 ± 1.1 |
| Time | Group | FFA (mM) | Glyc (mM) | TG (mM) |
|---|---|---|---|---|
| Baseline WT 1 | N | 0.29 ± 0.33 | 0.09 ± 0.09 | 1.10 ± 0.48 |
| P | 0.33 ± 0.28 | 0.08 ± 0.04 | 1.23 ± 0.78 | |
| Baseline ECT 1 | N | 0.29 ± 0.30 | 0.05 ± 0.04 | 1.14 ± 0.52 |
| P | 0.23 ± 0.15 | 0.05 ± 0.02 | 0.99 ± 0.61 | |
| Rest | N | 0.19 ± 0.16 | 0.06 ± 0.02 | 0.77 ± 0.30 * |
| P | 0.27 ± 0.19 | 0.07 ± 0.03 | 1.32 ± 0.59 | |
| 5th interval | N | 0.15 ± 0.06 | 0.08 ± 0.03 | 0.78 ± 0.35 |
| P | 0.18 ± 0.13 | 0.08 ± 0.02 | 1.49 ± 0.64 | |
| 20th interval | N | 0.16 ± 0.05 | 0.09 ± 0.02 | 0.83 ± 0.28 * |
| P | 0.20 ± 0.15 | 0.10 ± 0.02 | 1.32 ± 0.65 | |
| 35th interval | N | 0.18 ± 0.05 | 0.11 ± 0.04 | 0.74 ± 0.26 * |
| P | 0.20 ± 0.13 | 0.12 ± 0.03 | 1.47 ± 0.65 | |
| 45th interval | N | 0.20 ± 0.08 | 0.13 ± 0.06 | 0.70 ± 0.21 * |
| P | 0.24 ± 0.17 | 0.14 ± 0.05 | 1.49 ± 0.76 | |
| Cool-down | N | 0.23 ± 0.10 | 0.14 ± 0.07 | 0.69 ± 0.21 * |
| P | 0.26 ± 0.17 | 0.17 ± 0.05 | 1.32 ± 0.67 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blau, L.S.; Gerber, J.; Finkel, A.; Lützow, M.; Maassen, N.; Röhrich, M.A.; Hanff, E.; Tsikas, D.; Shushakov, V.; Jantz, M. Effects of Short-Term Sodium Nitrate versus Sodium Chloride Supplementation on Energy and Lipid Metabolism during High-Intensity Intermittent Exercise in Athletes. Appl. Sci. 2023, 13, 6849. https://doi.org/10.3390/app13116849
Blau LS, Gerber J, Finkel A, Lützow M, Maassen N, Röhrich MA, Hanff E, Tsikas D, Shushakov V, Jantz M. Effects of Short-Term Sodium Nitrate versus Sodium Chloride Supplementation on Energy and Lipid Metabolism during High-Intensity Intermittent Exercise in Athletes. Applied Sciences. 2023; 13(11):6849. https://doi.org/10.3390/app13116849
Chicago/Turabian StyleBlau, Larissa Sarah, Jan Gerber, Armin Finkel, Moritz Lützow, Norbert Maassen, Magdalena Aleksandra Röhrich, Erik Hanff, Dimitrios Tsikas, Vladimir Shushakov, and Mirja Jantz. 2023. "Effects of Short-Term Sodium Nitrate versus Sodium Chloride Supplementation on Energy and Lipid Metabolism during High-Intensity Intermittent Exercise in Athletes" Applied Sciences 13, no. 11: 6849. https://doi.org/10.3390/app13116849